Добро пожаловать в WordPress. Это ваша первая запись. Отредактируйте или удалите её, затем пишите!
Автор: aldrd
Реляционная модель данных
В 1970 году Эдгар Кодд опубликовал статью (Codd, 1970), в которой описал основы реляционной модели хранения данных. Практической реализацией этой модели стали все современные реляционные базы данных. Формализация модели привела к созданию реляционного исчисления и реляционной алгебры.
Основное элемент реляционной модели – это кортеж. Кортеж – это упорядоченный набор элементов, каждый из которых принадлежит определенному множеству или, иначе говоря, имеет свой тип. Совокупность однородных по структуре кортежей образует отношение.
Несколько более наглядно все это выглядит в терминах, используемых в базах данных (рисунок ниже). Отношение – это таблица с данными. Кортеж — строка таблицы. Какого типа кортежи содержатся в отношении, или, что то же самое, каков формат строк в таблице, определяется заголовком отношения или таблицы. Каждый из столбцов таблицы образует домен. Значения, которое могут принимать элементы домена, называются атрибутами. Строки таблицы – это совокупность атрибутов, соответствующих доменам.
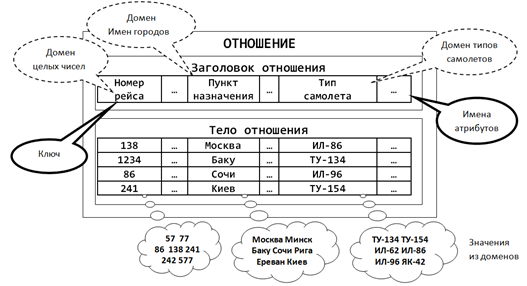
Пример отношения (Заборов)
Строки таблицы могут быть идентифицированы по своим атрибутам, то есть по тому, какие значения принимают элементы кортежа. Само содержание кортежа делает его непохожим на остальные. Но может так оказаться, что некоторые строки совпадут по своим атрибутам. Само по себе совпадение не страшно, но оно уже не позволяет использовать такой набор атрибутов, для однозначной идентификации кортежей в отношении. Чтобы идентификация была однозначной, вводят такое ключевое поле, которое для каждой строки принимает уникальное значение. Такой ключ может нести смысловую нагрузку, а может быть просто искусственно сгенерированным числом.
Совокупность всех отношений определяет базу данных. Каждое отношение хранит свою логическую часть информации. Чтобы получить определенные сведения может потребоваться сопоставление информации из разных отношений. Кодд описал восемь основных операций реляционной алгебры, позволяющих манипулировать с кортежами:
-
Объединение;
-
Пересечение;
-
Вычитание;
-
Декартово произведение;
-
Выборка;
-
Проекция;
-
Соединение;
-
Деление.
Замечательное свойство реляционной алгебры – это ее замкнутость, то есть операции над отношениями задаются таким образом, чтобы результат сам был отношением. То есть, имея несколько таблиц и производя соответствующие операции над ними, мы получим результатом тоже таблицу.
Смысл многих операций совпадает с соответствующими операциями из теории множеств. Общее представление об их сути дает рисунок ниже.
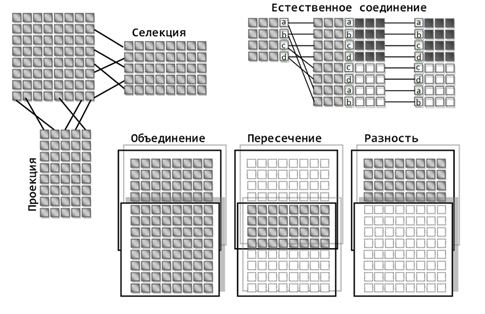
Пример операций над кортежами (Заборов)
Важно, что разные отношения могут содержать домены одного типа. Это значит, что если в двух кортежах встречаются одинаковые домены, внутри них одинаковые атрибуты, то можно говорить об определенной связи кортежей, содержащих эти атрибуты. Иначе говоря, если разные строки одной таблицы в одном из столбцов имеют одинаковые значения, то можно говорить об определенной связи этих строк. Или если в разных таблицах есть столбцы (домены) с одинаковым смыслом, то строки с одинаковыми значениями в этих столбцах оказываются связанными между собой.
Операция проекции позволяет получать отношения, состоящие из части элементов исходных отношений, ограничивая набор используемых доменов. Выборка или селекция позволяет получать отношения, содержащие только те кортежи, поля которых удовлетворяют условиям выборки. Например, можно выбрать только те кортежи, указанные домены которых имеют заданные значения атрибутов.
Совокупность всех операций над отношениями позволяет извлечь из базы данных любую интересующую информацию и сформировать ее в виде отношения (таблицы) с наперед заданными свойствами (заголовком).
Реляционной модель данных возникла не случайно, а явилась следствием необходимости оперировать с большими объемами разнообразных данных. Оказалось, что такая структура хранения данных и определенные в этой структуре операции удобны для решения широкого спектра прикладных задач. Можно предположить, что аналогичное удачное решение могла нащупать и природа в результате естественного отбора.
Описываемая нами система идентификаторов, понятий и событийной памяти во многом очень похожа на реляционную модель. Можно провести ряд аналогий:
-
Нейрон оперирует информацией с нескольких дендритных сегментов, каждый из которых настроен на данные определенного типа. Дендритные сегменты одного типа можно сопоставить с определенным доменом;
-
Сочетания понятий, которые описывают информацию, характерную для дендритного сегмента, соответствуют атрибутам, встречающимся в домене;
-
Понятия, используемые зоной коры, и идентификаторы, задающие структуру пакетов, характерную для этой зоны, определяют структуру доменов (заголовок);
-
Использование общих понятий при проекции информации между зонами соответствует использованию общих доменов в разных отношениях;
-
Совокупность зон коры, формирующих мозг, соответствует совокупности отношений, формирующих базу данных;
-
Ассоциативность, между воспоминаниями, соответствует связанности через общие атрибуты различных кортежей;
-
Распределенность воспоминания по зонам коры соответствует тому, как одно событие может породить несколько кортежей в разных отношениях, объединенных единым уникальным ключом;
-
Волна, описывающая текущее состояние мозга, может выступать аналогом запроса к базе данных. Так же, как результат операции над отношениями есть отношение, так и ответ мозга может быт совокупностью ассоциативно связанных описаний, совмещенных в одной волновой картине.
Конечно, между нашей моделью мозга и реляционными системами нет точного соответствия. Архитектура мозга значительно богаче, так как решает не только задачи хранения и извлечения данных, но и массу других совмещенных с этим функций. Однако даже имеющееся сходство позволяет лучше понять суть информационных процессов, происходящих в коре.
Кодирование сложных описаний
Вернемся к памяти и попробуем систематизировать, с какими типами информации, и, соответственно, типами описаний умеет оперировать наш мозг.
Первый тип – это простое описание, которое соответствует картине мгновенной активности коры. Это сочетание тех понятий, что детектированы мозгом именно сейчас.
Второй тип – это пакет простых описаний, соответствующих одному событию, одной мысли. В пакете несущественен порядок следования описаний. Перестановка слоев пакета не меняет общего смысла высказывания. Вспоминание пакета – это восстановление серии следующих друг за другом простых описаний.
Третий тип – это позиционное описание. В таком описании сохраняется связь одних объектов с другими, находящимися с ними в определенной системе отношений. Например, разновидность такого описания – это пространственное описание. Когда мы не просто фиксируем свое положение в пространстве, а увязываем его определенными описаниями с расположением других объектов.
Четвертый тип – это процедурное описание. Такое описание, в котором важна последовательность смены образов и сопутствующие этому интервалы. Например, восприятие речи определяется последовательностью звуков, при этом соотношение интервалов формирует интонацию, от которой сильно зависит общий смысл услышанной фразы. Вспоминание процедуры – это воспроизведение соответствующей последовательности образов.
И пятый тип – это хронологическое описание. Фиксация на продолжительных отрезках времени того в какой последовательности и с какими временными интервалами происходили те или иные события. Возможность вспомнить для хронологической памяти – это не воспроизведение сразу всего, относящегося к одной хронологии, а возможность перейти от одного описания к другому, связанному с ним общей временной последовательностью.
Несложно заметить, что многие описания так или иначе завязаны на время. Пакетное описание – это серия следующих друг за другом образов. Процедурное описание учитывает последовательность событий. Хронологическое описание требует учета позиционирования событий во времени.
Такая зависимость описаний от времени послужила поводом к возникновению соответствующих моделей. Наиболее известная из них – это пропагандируемая Джефом Хокинсом концепция иерархической темпоральной памяти (HTM) (Хокинс, 2011). Он и его коллеги исходят из того, что темпоральная смена событий – это единственное, что позволяет связать между собой отдельные информационные образы. Из этого делается вывод, что базовый информационный элемент коры должен работать не со статичными образами, а с временной последовательностью. В концепции HTM элемент хранения информации – это развернутая во времени последовательность сигналов. Узнавание – это определение совпадения двух последовательностей. При этом особый акцент делается на способность HTM к предсказанию. Как только нейрон узнает начало знакомой ему последовательности, он становится способен по своему опыту предсказать запомненное им продолжение. Описание текущей картины в HTM – это активность тех нейронов, которые откликнулись на текущую смену событий.
Сложности такого подхода достаточно очевидны. Во-первых, требование соблюдения временных масштабов. Небольшое ускорение или задержка в поступлении данных могут нарушить алгоритм узнавания. Во-вторых, необходимость переводить все статичные образы во временные последовательности перед тем, как кора сможет с ними оперировать. И тому подобное.
В нашей модели система идентификаторов дает нам универсальный инструмент, который одинаково хорошо подходит для описания всех возможных типов памяти. Основная идея проста – каждое простое описание представляет собой составной идентификатор, содержащий в себе все необходимое для указания всего набора, как ассоциативных, так и временных взаимосвязей.
На рисунке ниже приведено условное изображение такого простого описания. Простое описание – это волна, несущая в себе несколько наборов идентификаторов разных типов. Основное содержание кодируется набором идентификаторов понятий, описывающих суть происходящего. Идентификатор слоя помечает основное содержание, отделяя его от остальных простых описаний. Идентификатор пакета объединяет несколько слоев, относящихся к одному сложному описанию. Идентификаторы места, времени и последовательности создают систему соответствующих связей между сложными описаниями.
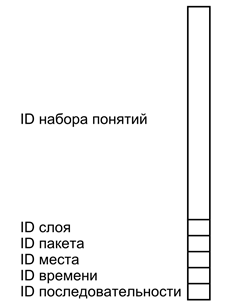
Формат простого описания
Возьмем предыдущий пример и обозначим волны идентификаторов, соответствующие используемым понятиям (concepts), как C1…C7 (рисунок ниже).
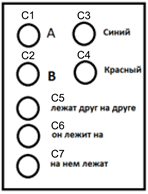
Понятия, используемые для описания
Тогда описание того, что «красный объект A лежит на синем объекте B» будет выглядеть, как показано на рисунке ниже.
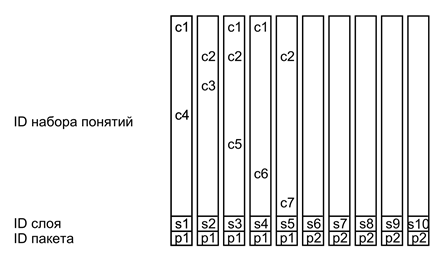
Пример сложного описания
В этом примере каждый из слоев пакета является простым описанием со своим идентификатором слоя . Все слои пакета имеют общий идентификатор пакета p1. Когда заканчивается одно сложное описание, другое, следующее за ним имеет отличный от него идентификатор пакета p2 (понятия второго описания на рисунке не показаны).
Чтобы такая конструкция была работоспособной, мозгу необходима достаточно сложная система, создающая идентификаторы, формирующие пакеты. Причем для каждой из зон коры может потребоваться свой набор таких идентификаторов, уместный именно для нее.
Например, возьмем последовательность зрительного восприятия. Скачкообразные микродвижения глаз, называемые микросаккадами, заставляют глаз сканировать малый фрагмент изображения, приходящийся на центр сетчатки. Все образы, которые получаются в процессе такого сканирования, предположительно могут объединяться общим идентификатором. Микродвижениями глаз управляют верхние бугорки четверохолмия. Можно предположить, что именно они кодируют такой идентификатор. После нескольких микросаккад происходит сильный прыжок, называемый саккадой (выше на рисунке с головой Нефертити показаны именно саккады). Каждая саккада вызывают смену идентификатора микросаккад.
Можно предположить, что микросаккады принципиально важны для первичной зрительной коры. Общий идентификатор сообщает коре, что сери подряд идущих образов описывает один и тот же объект, но в разных его позициях на сетчатке, что позволяет объединить их в единое описание и реализовать инвариантное к положению на сетчатке узнавание.
Более продолжительное событие — серия саккад. Так как серия относится к разглядыванию единой картины, то получаемые описания тоже можно связать между собой еще одним общим идентификатором – идентификаторам саккад. Но этот идентификатор существенен уже не для первичной, а для вторичной и более глубоких уровней зрительной коры, где происходит последующая обработка информации. Идентификатор, сообщающий коре, что все, что мы видим во время серии саккад – это одна и та же картина, позволяет соотнести между собой одни и те же образы, видимые разными местами сетчатки.
Смена идентификатора саккад должна происходить, когда существенно меняется разглядываемая картина. Например, при сильном повороте головы, переключении внимания, смене плана или сцены в кино. Переключение внимания может кодироваться элементами лимбической системы мозга и распространяться на множество, завязанных на это, зон коры. Одновременно с этим в системе описаний присутствуют идентификаторы гиппокампа, кодирующие пространственно-временные описания событий. Короче, система идентификаторов, определяющих пакет, может быть достаточно сложна, и определяться особенностями той информации, с которой имеет дело каждая конкретная зона коры.
Используя идентификаторы несложно организовать фиксацию последовательности событий. Например, если взять идентификатор, состоящий из двух фрагментов, то поочередно меняя по одному из них, можно получить ассоциативную связанность соседних описаний (рисунок ниже).
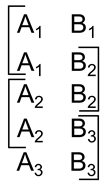
Кодирование последовательности
Каждый такой идентификатор будет содержать элемент от предыдущего и последующего идентификатора. Запомнив временную последовательность образов с такими идентификаторами, мы для каждого образа сможем найти его двух соседей по временной шкале. Несложно усложнив идентификатор можно закодировать не только общую связанность, но и направление течения времени.
Надо отметить, что в нашей модели каждое воспоминание имеет богатую систему идентификаторов. Это позволяет получить доступ к воспоминанию через множество совершенно различных ассоциаций. Можно вспомнить что-либо, исходя из совпадения смыла описаний. Можно проассоциировать информационные картины по месту или времени описываемых событий. Можно воспроизвести последовательность образов, относящихся к одному событию. Нетрудно заметить, что такой доступ к воспоминаниям имеет много общего с подходами, которые используются при создании традиционных реляционных баз данных.
Пакетное представление информации
Информация, которой оперирует мозг, должна, с одной стороны, достаточно полно описывать происходящее, с другой стороны, должна храниться так, чтобы допускать выполнение над собой требуемых мозгу операций. В принципе, формат описания информации и алгоритмы ее обработки – вещи тесно связанные между собой. Первое во многом определяет второе. Поэтому говоря о том, как могут быть организованы данные, хранимые мозгом, мы, хотим того или нет, во многом предопределяем систему последующих мыслительных процессов. Так как разговор о принципах мышления нам предстоит позже, то сейчас мы сделаем акцент только на том, как обеспечить полноту текущего описания и последующего хранения информации. При этом подразумевая, что если, дойдя до мышления, окажется, что выбранный нами формат данных подошел под требуемые алгоритмы, то значит, нам повезло и мы пошли по правильному пути.
Чтобы понять, какой формат описаний использует мозг, проследим последовательность зрительного восприятия. Разглядывая изображение, мы «сканируем» его быстрыми движениями глаз, называемыми саккадами (рисунок ниже). Каждая из них помещает в центр зрения один из фрагментов общей картины. На зонах зрительной коры возникают описания, соответствующие тому, что мы видим в этот момент в центре, что видит периферия и каково смещение в результате только что проделанной саккады. Каждая следующая саккада порождает новую картину. Эти описания сменяют друг друга одно за другим.
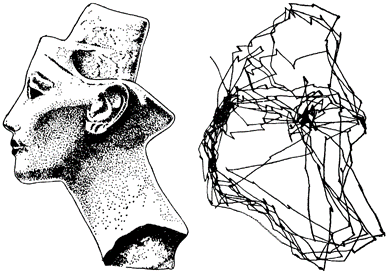
Запись движений глаз при свободном разглядывании в течение двух минут (Ярбус, 1965)
Так, разглядывая лицо, мы сначала, например, четко видим и распознаем один глаз, тот на который направлен взгляд. Остальные элементы лица, попадающие на относительную периферию зрения — нос, рот и так далее, мы узнаем с меньшей, но то же достаточно высокой вероятностью. После каждой саккады центральный фрагмент меняется, но общий набор узнанных элементов сохраняется неизменным.
В принципе, каждого из таких отдельных описаний, возникающих между саккадами, достаточно чтобы сказать, что перед нами лицо и даже узнать, кому оно принадлежит. Но каждое отдельное описание достоверно говорит только о том объекте, который для него расположен по направлению взгляда. Остальные объекты определяются достаточно приблизительно.
Если мы захотим получить более полное и подробное представление о лице, то для этого подойдет совокупность всех описаний, которые возникнут во время сканирования. При этом важно будет не только описание того, что за объекты узнаны, но и информация об сопутствующих смещениях взгляда. И тут мы подходим к очень важному моменту. Что же является тем итоговым описанием, которое должен выдавать зрительный анализатор? Просто картина активности ряда понятий? Это соответствует только той части описания, что мы видим прямо сейчас. А как же остальное? Получается, что корректное, не теряющее информации, описание – это пакет следующих друг за другом более простых описаний. Где каждый из слоев такого временного пакета описывает только некоторую часть информации, а полное описание получается, как их совокупность. Это справедливо при условии, что все описания в пакете соответствуют одному событию, то есть, получены до глобального переключения нашего внимания.
Если взять моментальный снимок активности коры мозга, то описание происходящего можно сопоставить с перечислением активных понятий на каждой из ее зон. Но такое описание имеет существенный недостаток. Предположим, что мы хотим описать натюрморт, изображенный на рисунке ниже.
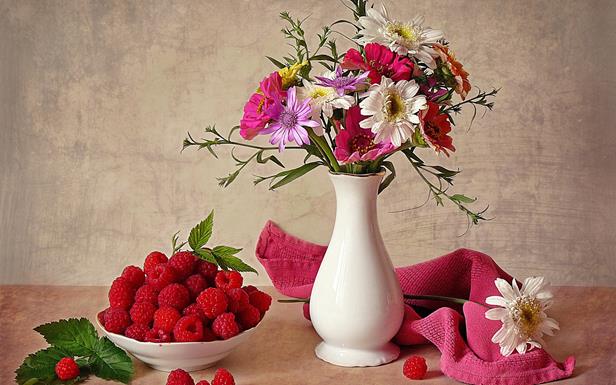
Мы можем это сделать, например, так:
-
Ваза чуть правее центра;
-
Букет в вазе;
-
Полотенце справа от вазы;
-
Белый цветок на полотенце;
-
Миска с малиной слева;
-
Малина на листе слева от миски;
-
Три малины перед миской;
-
Малина справа от вазы.
Общее описание складывается из набора таких коротких описаний. Каждое короткое описание можно, с некоторыми оговорками, заменить перечислением понятий, входящих в него. Но если мы захотим собрать итоговое описание, просто сложив все понятия, участвующие в коротких перечислениях, то нас постигнет неудача. При сложении исчезнет часть информации, так как станет непонятно, что к чему относится. Но, кроме того, окажется, что надо некоторые понятия использовать несколько раз. Например, малина она и слева, и справа, и перед миской. А если мы хотим использовать это «просто собранное» описание как аналогию того, как описывается такой натюрморт на зонах коры, то окажется, что обобщение «малина» у нас одно и оно не может одновременно быть «активным три раза». Выход из этого положения, который мне видится вполне логичным, — это использовать пакетное описание. Каждое простое описание может складываться из банального перечисления активных понятий. Полное же описание получается, как набор простых описаний. Поскольку простые описания разнесены между собой по времени, то, с одной стороны, понятно, что к чему относится, а, с другой стороны, одно и то же понятие может возникать несколько раз в разных слоях пакета в разных контекстах.
Такое пакетное представление очень хорошо соотносится с рассуждениями относительно объема внимания человека. Психологи, изучая свойства внимания, установили, что есть предел того количества объектов, на которых человек может одновременно сконцентрироваться. Обычно этот предел не превышает семи объектов. Первым измерения объема внимания с использованием механического тахистоскопа сделал основатель экспериментальной психологии Вильгельм Вундт.

Тахитоскоп – прибор, с помощью которого можно предъявлять последовательные зрительные стимулы
Оценить объем внимания очень просто. Посмотрите на предыдущий натюрморт и попробуйте сосчитать, сколько отдельных элементов вы способны, нет, не запомнить, это другое, а удержать в голове одновременно. Или возьмите семизначный телефонный номер, например, 1145618 и попробуйте «удержать» его в голове. Скорее всего, чтобы он не пропал, вам придется зациклено повторять его про себя. Если цифр в номер будет больше семи, что велик шанс, что удержать их все в памяти не удастся. Предельное количество воспринимаемых одновременно объектов натюрморта или цифр номера дает оценку объему вашего внимания.
Сделанное нами предположение о пакетном представлении информации в коре мозга позволяет сопоставить каждый из объектов, удерживаемых во внимании, с одним из слоев информационного пакета.
Если представить кору, состоящую из небольшого количества понятий, способную формулировать совсем простые мысли относительно двух объектов «A» и «B», то пакет, соответствующий мысли: «красный объект A лежит на синем объекте B», будет выглядеть, как показано на рисунке ниже.
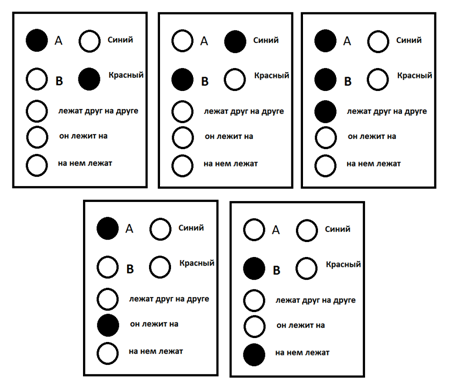
Пример информационного пакета
Консолидация памяти
Попробуем представить информационную емкость мозга. Так как в основе памяти лежат нейроны, то для начала уточним, с каким количеством нейронов мы имеем дело. Наиболее точная оценка на сегодня, принадлежит Сюзанне Херкулано-Хузель (Frederico A.C. Azevedo, Ludmila R.B. Carvalho, Lea T. Grinberg, José Marcelo Farfel, Renata E.L. Ferretti, Renata E.P. Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009). По этой оценке на кору приходится 82% общей массы мозга и 19% от общего числа нейронов (рисунок ниже). По этой оценке кора состоит из порядка 16 миллиардов нейронов (1.6×1010).
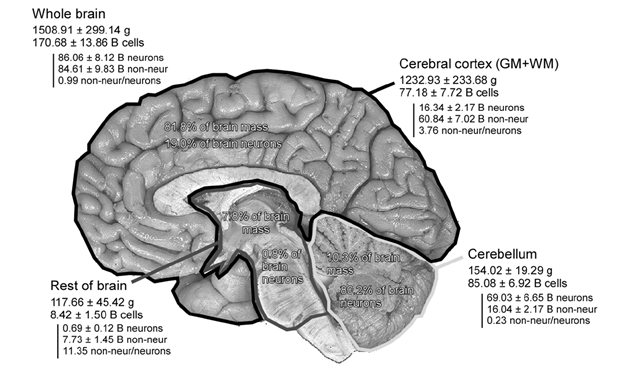
Количество нейронов в различных частях мозга (в миллиардах) (Frederico A.C. Azevedo, Ludmila R.B. Carvalho, Lea T. Grinberg, José Marcelo Farfel, Renata E.L. Ferretti, Renata E.P. Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009)
В моделях, использующих пластичность синапсов для объяснения природы памяти, емкость памяти считается достаточно просто. Характерное для нейрона количество синапсов – 10 000 (104). Если полагать, что один синапс хранит один бит информации, то получается 1.6×1010 x 104 = 1.6×1014 бит или приблизительно 2×1013 байт. Это около 20 000 гигабайт или 20 терабайт. Объем достаточно скромный даже по сегодняшним меркам.
Если предположить, что активная жизнь человека около 80 лет, из которых он треть проводит во сне, то это составит 80x365x(24-8)x60x60 = 1681920000 (1.7×109) секунд. Получается, что если не учитывать организацию памяти, а просто заполнять ее в течение жизни, то можно позволить скорость записи 2×1013/1.7×109=1.18×104 байт/секунду или около 10 кбайт/секунду.
В нашей модели оценка объема памяти выглядит несколько иначе. Оценка синаптической памяти не меняется, те же 20 терабайт. Но это только память, хранящая образы нейронов-детекторов. Основные объемы приходятся на внесинаптические рецептивные кластеры. Если задаться оценкой, что один внесинаптический образ формируется из сигналов порядка 100 синапсов, то можно оценить информационную емкость одного элемента, как 100 бит. Исходя из оценки, что поверхность нейрона может содержать порядка 100 000 рецептивных кластеров, получаем емкость памяти 1.6×1010x105x100 = 1.6 x1017 бит или 2×1016 байт. Что составляет 20 000 терабайт или 20 петабайт.
Если предположить, что половина этой памяти уходит на способность коры распространять волны идентификаторов, то объем событийной памяти можно оценить в 10 петабайт. Этот объем вызывает определенное уважение. Исходя из него, скорость записи информации при непрерывном запоминании может составлять 1016/1.7×109=5.88×106 байт/секунду или около 6 мегабайт/секунду. Если предположить, что фиксация энграмм происходит несколько раз в секунду, то можно дать оценку информационной емкости одной энграммы в районе 1 мегабайта.
Потенциально, наша память способна хранить достаточно детальное описание всего, что случается с нами в жизни. Тотальность памяти подтверждается наличием людей-феноменов, которые действительно «помнят все». Александр Лурия в результате многолетних наблюдений задокументировал один из примеров такой безграничной памяти:
«Оказалось, что память Ш. не имеет ясных границ не только в своем объеме, но и в прочности удержания следов. Опыты показали, что он с успехом – и без заметного труда – может воспроизводить любой длинный ряд слов, данных ему неделю, месяц, год, много лет назад. Некоторые из таких опытов, неизменно кончавшихся успехом, были проведены спустя 15 – 16 лет (!) после первичного запоминания ряда и без всякого предупреждения. В подобных случаях Ш. садился, закрывал глаза, делал паузу, а затем говорил: «да-да… это было у вас на той квартире… вы сидели за столом, а я на качалке… вы были в сером костюме и смотрели на меня так… вот… я вижу, что вы мне говорили…» – и дальше следовало безошибочное воспроизведение прочитанного ряда» (Лурия, 1968).
Но как бы не была безгранична наша память, хорошо известен тот факт, что гораздо легче и детальнее нам вспоминаются недавние события. Еще в 1885 году психологом Германом Эббингаузом была получена кривая забывания (Ebbinghaus, 1885)(рисунок ниже).
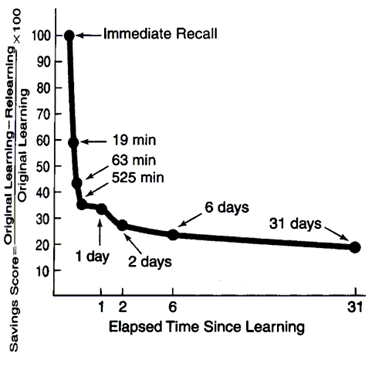
Кривая забывания
Наиболее резкое забывание наблюдается в первые минуты, затем процесс забывания замедляется. Через 30 дней память выходит на стабильную фазу, когда дальнейшего забывания не происходит. В своих опытах Эббингауз использовал наборы бессмысленных трехбуквенных слов, которые заучивал наизусть. Заучивание несколько отличается от простого восприятия, то есть кривая забывания для единожды услышанной фразы или увиденного образа будет несколько иная.
Запомнить что-либо более сильно позволяет повторение. Повторяя материал, мы не просто освежаем воспоминания и начинаем новый цикл забывания. После повторения, асимптота, на которую выходит кривая забывания, поднимается существенно выше по сравнению со своим предыдущим уровнем (рисунок ниже). Каждое новое повторение повышает шансы на то, что, спустя продолжительное время, нам удастся все относительно безошибочно вспомнить.
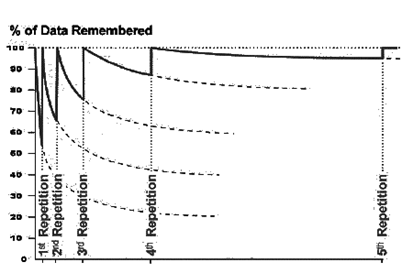
Кривая забывание при повторениях
Важно отметить, что повторение не обязательно должно сводиться к еще одному предъявлению заучиваемого образа. Воспоминание, достаточно точно воспроизводящее запомненное событие, так же является повторением. То есть чем чаще мы вспоминаем о чем-либо, тем прочнее оно врезается в нашу память.
Традиционная трактовка таких свойств памяти опирается на представление о переводе воспоминаний из кратковременной памяти в память долговременную. При этом часто, опираясь на компьютерные аналогии хранения данных, подразумевают перемещение информационных следов из одних частей мозга в другие. Так принято считать, что гиппокамп, работая наподобие оперативной памяти компьютера, накапливает текущие воспоминания, а затем перемещает их на кору, аналогично тому, как происходит сохранение компьютерных данных на долговременных носителях.
Процесс перехода воспоминаний из краткосрочной в долгосрочную форму принято называть консолидацией памяти. Это объективный и достаточно хорошо изученный процесс. Кроме того, что мы можем наблюдать изменения памяти на собственном опыте, есть и прямые подтверждения наличия консолидации. Так известно, что травмы головы, электрошок, судорога нарушают нормальный ход формирования памяти. Они вызывают ретроградную амнезию, когда воспоминания, относящиеся к некоторому промежутку времени, предшествующему сбою, оказываются утеряны.
Аналогичный эффект потери памяти получается при введении в мозг веществ, блокирующих экспрессию определенных генов, производящих белки, участвующие в работе памяти.
Кроме того, эффект ретроградной амнезии знаком многим по существенной потере памяти после чрезмерного употребления алкоголя. Определенная критическая доза выпитого спиртного настолько нарушает процесс консолидации, что часть воспоминаний полностью стирается, создавая невосполнимый провал в памяти. При этом происходит не потеря способности к текущему запоминанию, как при нарушениях в работе гиппокампа, а именно стирание уже полученных воспоминаний, точнее не перевод их в долговременную фазу хранения. Больной с синдромом Корсакова теряет воспоминание, как только происходит переключение его внимания. Человек, выпивший критическую дозу, все равно помнит недавние события, и во многих случаях может поддерживать связанную беседу. Однако наутро все воспоминания, начиная с определенного момента, оказываются стертыми.
Для объяснения эффекта консолидации необязательно замыкаться на компьютерной аналогии, связанной с физическим перемещением данных. В нашей модели консолидацию и ее смысл можно описать следующим образом. Как мы говорили ранее, внесинаптические метаботропные рецептивные кластеры обладают эффектом гистерезиса. Это значит, что если нейрон, генерирует спайк, то скачек мембранного потенциала при выполнении ряда условий, описанных нами ранее, может послужить поводом для перехода рецептивного кластера в устойчивое состояние. Устойчивое состояние – это либо состояние, в котором концевые части рецепторов вытолкнуты из мембраны и рецептор приобретает чувствительность к определенному сигналу, либо состояние их утопленности в котором рецептор, наоборот чувствительность теряет. Такое изменение происходит с рецептивными кластерами, для которых выполнилось условие интерференции медиаторов от окружающих их синапсов, то есть которые настроились на определенный образ.
Переход рецептивных кластеров в такое состояние происходит одновременно у огромного количества нейронов, активных в текущий момент времени. Что означает, что этот текущий момент запоминается с чрезвычайной избыточностью и детализацией. Это ведет к тому, что при прочих равных, текущие события имеют значительно более высокую вероятность быть вспомненными по сравнению с событиями более отдаленными во времени.
Такую гиперподробную память можно назвать оперативной или кратковременной памятью, но не стоит путать ее с текущей вызванной и волновой активностью нейронов коры. Активность нейронов формирует описание происходящего, кратковременная же память хранит картины таких относительно недавних состояний. Причем кратковременная память ничем не отличается от остальной памяти, за исключением того, что свежие воспоминания более детальны, так как используют существенно большее количество рецептивных кластеров, чем остальные энграммы.
Продолжительность сверхоперативной памяти – секунды. Затем большая часть внесинаптических рецепторов возвращается в исходное состояние, обретая готовность участвовать в новых актах запоминания. Но существенная часть рецептивных кластеров по тем или иным причинам остается в состоянии фиксации образа. Они переходят в следующую устойчивую фазу, которая позволяет им существовать дальше и соответственно хранить фрагмент своего воспоминания.
Можно предположить, что каждое воспоминание проходит несколько стадий консолидации, лишаясь на каждом этапе части своих рецептивных кластеров. Самый простой алгоритм забывания – вероятностный. Регулируя вероятность прохождения рецептивным кластером очередного этапа консолидации, можно получить требуемый объем для каждого воспоминания к моменту его перехода в долговременную память.
Фазы консолидации сопровождается синтезом советующих белков, которые определяют общее течение всех процессов. Ход консолидации для метаботропных и для ионотропных рецепторов имеет свои особенности, но достаточно похож идеологически. Сразу после обучения в ядре нейрона начинается экспрессия немедленных «ранних генов». Эти гены кодируют транскрипционные факторы. Транскрипционные факторы запускают транскрипцию «поздних генов». Примерный график такой процедуры показан на рисунке ниже.
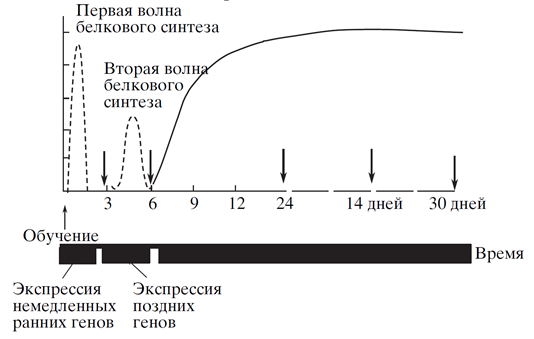
Экспрессия «ранних» и «поздних» генов (Е.В. Муравьева, К.В. Анохин, 2006).
Известны разные белки, относительно которых доказано их участие в процессе формирования памяти. Arc,
CPEB, Kibra – это только малая часть из них, хотя и наиболее часто упоминаемая. Надо помнить, что информация оставляет на нейроне два типа следов. Один связан с изменением чувствительности его синапсов, другой с фиксацией изменений во внесинаптических рецепторах. Первое влияет на свойства нейрона, как детектора. Второе обеспечивает способность нейронов распространять волны идентификаторов, хранить событийную память, формировать ассоциативность и тому подобное. Как и в какой последовательности происходит белковая регуляция этих процессов вопрос не простой и пока не имеющий общепринятого ответа, поскольку нет пока и общепризнанной модели памяти. Поэтому в нашей модели мы будем делать акцент на общую идеологию процессов, к сожалению, не имя возможности доказательно описать их белковую регуляцию.
Консолидация фактически задает таймер, который отсчитывает стадию жизни элементов воспоминания и регулирует вероятность их перехода в следующую фазу. Можно предположить, что повторение опыта или что равносильно воспоминание о нем, изменяют ход консолидации, повышая шансы рецептивных кластеров на переход в фазу долговременной памяти. То есть чем больше повторений или чем чаще воспоминания, тем более значимый след оставляет событие. И тем выше вероятность его восстановления при соответствующем информационном запросе.
Так как события эмоционально значимые заставляют нас чаще вспоминать о них, по сравнению с событиями нейтральными, то это может быть одной из причин, почему эмоционально окрашенные события формируют более прочные и устойчивые следы в нашей памяти.
Но воспоминания о пережитом опыте не так просты, как кажутся. Психологам давно известен феномен трансформации воспоминаний. Первым его показал еще в 1932 году Фредерик Бартлетт (Bartlett, 1932). Он установил, что если человек, вспоминая определенное событие, добавляет какие-либо детали, то исходное воспоминание замещается новым, а измененные детали становятся частью этого нового воспоминания. Причем человек не осознает этих изменений, а считает, что все было именно так как он помнит теперь. Такое изменение воспоминаний принято называть реконсолидацией памяти.
Реконсолидация памяти хорошо вписывается в нашу модель. Воспоминание о каком-либо событии сопровождается восстановлением его уникального идентификатора. Если по какой-либо причине происходит искажение исходного воспоминания, то текущий информационный образ запоминается с тем же самым идентификатором, что и основное воспоминание, так как именно этот идентификатор присутствует в пространстве коры. А значит, неизбежно происходит описанный Барлеттом феномен трансформации.
Есть очень интересные опыты, которые показывают, что реконсолидация может сопровождаться не только трансформацией, но и стиранием памяти. Но поскольку эти опыты больше относятся к поведению, чем к событийной памяти мы рассмотрим их позже в соответствующем разделе.
Общую идеологию событийной памяти можно описать следующим образом. Поступающая информация создает свежие воспоминания с огромной избыточностью и детализацией. Это делает их исключительно доступными для оперативного доступа. Такая память живет не долго. Она память подвергается стадиям консолидации, каждая из которых уменьшает количество рецептивных кластеров, формирующих энграммы. В результате всех этапов консолидации, продолжающихся, по-видимому, около месяца, энграммы приобретают стабильный объем и переходят в фазу долговременного хранения.
Ранее мы оценили порядок объема событийной памяти мозга в 10 петабайт. Откуда цыганским методом вывели оценку размера энграммы в 1 мегабайт. Эта оценка может быть справедлива для старых, прошедших все стадии консолидации, воспоминаний. Большая часть информации, с которой мы оперируем, относится к текущему дню. Для этой информации обеспечивается развернутое избыточное хранение. Можно предположить, что такая актуальная память занимает существенный объем от общей памяти мозга. Если бы я был конструктором человеческого мозга, то я бы эмпирически отвел на свежие воспоминания половину всей доступной памяти, а оставшуюся половину определил под старые воспоминания.
При таком эмпирическом предположении скорость записи информации мозгом можно оценить, как 5×1015/(16x60x60), что составляет около 1011 байт или 100 гигабайт в секунду. Конечно, это значительно больше, чем весь объем информации, поступающий от всех вместе органов чувств. Но разговор идет не о входном объеме, а о развернутом описании, состоящем из множества описаний, возникающих на каждой из зон коры.
В такой прикидке начальный размер одной энграммы составит порядка 10 гигабайт. Собственно, процесс консолидации памяти – это постепенное сведение этого чрезвычайно избыточного объема до уровня порядка 1 мегабайта.
Тем не менее, каким бы на самом деле не было соотношение объемов новой и старой памяти, природа должна была оптимизировать его так, чтобы не допустить неоправданного резервирования простаивающих объемов. Это значит, что с учетом крайне высокой скорости записи текущей информации, может наступить момент переполнения всей доступной памяти. Можно предположить, что сонливость – это и есть состояние, когда информационные ресурсы мозга подходят к своему заполнению, и требуется расчистка памяти.
В принципе, оперативная консолидация памяти происходит постоянно, непрерывно сжимая объемы энграмм, мозг обеспечивает пространство для новых воспоминаний. Но можно предположить, что оперативная консолидация имеет ряд недостатков. Так в нашей модели нейроны-детекторы образуют паттерны, соответствующие понятиям. Каждое понятие образуется множеством паттернов, расположенных в разных местах коры. Чтобы паттерн запустил волну, должна активироваться существенная часть нейронов паттерна. Это значит, что если пользоваться вероятностной консолидацией, просто удаляя часть рецептивных кластеров, то мы постепенно разрушим все энграммы до такого уровня, когда ни один из паттернов не будет в состоянии излучить волну, связанную с этим воспоминанием. Чтобы избежать этого, разумно использовать выборочную консолидацию.
Выборочная консолидация предполагает, что мы избирательно определяем на каких паттернах стоит оставить четкие следы воспоминаний, а на каких стоит их полностью удалить. Тогда при том же объеме энграммы и сохранении ее распределенного характера удастся избежать малой плотности энграмм на отдельных паттернах.
Можно предположить, что выгоднее использовать для сохранения энграмм те паттерны, которые своим положением на коре наиболее точно соответствуют активности, связанной с общим контекстом воспоминания. Но процедура определения таких паттернов требует повторного технического воспроизведения запомненных событий. Однако такое воспроизведение несовместимо с нормальной работой мозга, так как, протекая одновременно с текущим восприятием, будет восприниматься мозгом как галлюцинация. Уместно предположить, что оптимальная консолидация, связанная с удалением большей части, накопленной за день информации, не возможна или затруднена при обычной работе мозга. Тогда получается, что сон – это именно то состояние, которое необходимо мозгу для выполнения процедур подобного рода.
Кроме того, что мозг накапливает событийные воспоминания, одновременно с этим происходит обучение нейронов, как детекторов. И то и другое основано на изменении структуры нейронных рецепторов. Изменения в рецепторах управляются продуктами экспрессии определенных генов. Продукты экспрессии генов – это белки. Часть из них непосредственно участвует в клеточных процессах, часть является транскрипционными факторами, которые регулируют последующую экспрессию генов. По изменению концентрации соответствующих белков можно судить о течении процессов, связанных с обоими видами консолидации.
Увидеть и оценить концентрацию белков можно через окрашивание среза мозговой ткани красителем, избирательно реагирующим на требуемый белок. Подобные наблюдения показали, что наиболее масштабные изменения происходят именно во время сна (Chiara Cirelli, Giulio Tononi, 1998) (Cirelli, 2002) (рисунки ниже).

Распределение белка Arc в теменной коре крысы после трех часов сна (S) и после трех часов спонтанного бодрствования (W) (Cirelli, 2002)

Распределение транскрипционного фактора P-CREB в корональных участках теменной коры крысы после трех часов сна (S) и в случае лишения сна на три часа (SD) (Cirelli, 2002)
Консолидация памяти во сне происходит в несколько этапов. У здорового человека сон начинается с первой стадии медленного сна, которая длится 5-10 минут. Затем наступает вторая стадия, которая продолжается около 20 минут. Еще 30-45 минут приходится на периоды третей и четвертой стадий. После этого спящий снова возвращается во вторую стадию медленного сна, после которой возникает первый эпизод быстрого сна, который имеет короткую продолжительность — около 5 минут. Во время быстрого сна глазные яблоки очень часто и периодически совершают быстрые движения под сомкнутыми веками. Если в это время разбудить спящего, то в 90% случаев можно услышать рассказ о ярком сновидении. Вся эта последовательность называется циклом. Первый цикл имеет длительность 90-100 минут. Затем циклы повторяются, при этом уменьшается доля медленного сна и постепенно нарастает доля быстрого сна, последний эпизод которого в отдельных случаях может достигать 1 часа. В среднем при полноценном здоровом сне отмечается пять полных циклов.
Стадии сна можно объяснить необходимостью обработать информацию различного типа. Алгоритмы консолидации событийной памяти, волновой памяти, связанной с распространением идентификаторов, и памяти, связанной с пластичностью синапсов, могут сильно различаться.
В наших предположениях хорошо объясняется цикличность сна. Полная консолидация, как каждому известно из личного опыта, требует около восьми часов. Но обстоятельства могут заставить нас проснуться раньше. Если бы восьмичасовой сон состоял из одного цикла, разделенного на несколько фаз, то преждевременное прерывание сна вело бы к тому, что начальные фазы сна успевали бы завершиться полностью, при том, что поздние еще даже не начинались. То есть часть информации оставалась бы совершенно необработанной, при этом полностью исчерпав за предыдущий день соответствующий ресурс мозга. То есть мозг, по сути, был бы в неработоспособном состоянии. Несколько циклов позволяют при преждевременном пробуждении хотя бы частично выполнить каждую из процедур консолидации и привести мозг в адекватное состояние. Но потребность в полноценной консолидации не исчезает, и впоследствии мозг должен будет компенсировать упущенные циклы более продолжительным сном, что обычно называется «отоспаться».
Похоже, что консолидация памяти связана с фазами медленного сна. По мере ее завершения эти фазы становятся все короче, уступая место быстрому сну. Это объясняет тот факт, что при полном завершении консолидации, завершающий этап сна состоит исключительно из продолжительной фазы быстрого сна.
Быстрый сон – это те самые сновидения, которые образуют причудливые фантазии, только отдаленно связанные с реальным опытом. Закономерность, подмеченная для сновидений еще Зигмундом Фрейдом, гласит, что во сне проступают явления, наиболее беспокоящие нас, важные для нас, концентрирующие суть наших терзаний и переживаний (Фрейд, 1900). В большинстве случаев сны очень быстро забываются. Мы видим сны каждую ночь, но стремительность, с которой мы их забываем, заставляет многих считать, что они вообще не видят снов.
В нашей модели стирание сновидений реализуется достаточно просто. Для этого достаточно сделать так, чтобы следы идентификаторов гиппокампа не фиксировались надолго, а подвергались быстрому расформированию. Это приведет к тому, что мы будем безвозвратно забывать о сновидениях. Но если, проснувшись, мы успеем вспомнить о недавнем сне, то возникнет новое уже традиционное воспоминание, которое сохранит события сна.
Можно предположить, что для снов стирание следов идентификаторов гиппокампа не сопровождается удалением ассоциативных следов. Тогда смысл сновидений можно объяснить усилением системы ассоциаций для наиболее значимых и важных для нас понятий. Что в свою очередь способствует последующему восприятию мира таким образом, что именно эти понятия будут наиболее акцентированы в системе описания новых событий.
Всем хорошо известно, что «утро вечера мудренее». Но в чем суть этой мудрости? Иногда утренние озарения объясняют тем, что де, мол, мозг ночью решает дневные задачи. К такому объяснению подталкивает, во-первых, тот факт, что зафиксированная во многих исследованиях ночная активность мозга сильно напоминает активность пережитого днем, а, во-вторых, созвучность сновидений тем темам, что нас тревожат наяву. Но уместней предположить, что мозг не решает ночью дневные задачи, он работает над оптимизацией системы понятий и воспоминаний. Утром эта оптимизация позволяет значительно легче найти объяснения вчера еще непонятным вопросам. Чтобы правильно задать вопрос, надо знать большую часть ответа. Та работа, которую делает ночью мозг — это и есть подготовка той самой большей часть ответа — возможности подойти к вопросу с «правильным» набором внутренних понятий и избавившись от излишней детальности ненужных воспоминаний. То есть, если все, сказанное сейчас про сон, кажется вам слишком туманным, то попробуйте вернуться к этому завтра.
Нет смысла жалеть о времени, проведенном во сне. Сон – это замечательная возможность «поумнеть», не прикладывая никаких усилий. Единственное, что все-таки необходимо – это предварительно загрузить голову достаточным объемом свежей информации. Причем этой информацией могут быть не только новые факты, но и старые знания, заново вспомненные или обдуманные. Любые наши размышления оставляют след свежих воспоминаний на коре. Во сне происходит консолидация новых размышлений и старого опыта, что дает возможность возникнуть более глубоким обобщениям. Это означает, что дневные размышления по какому-либо вопросу имеют смысл, даже если они не приводят к немедленному ответу. Эти размышления создают те свежие воспоминания, относящиеся к интересующей нас теме, что ночью дадут мозгу возможность поработать над оптимизацией всей системы наших понятий. Можно вспомнить о том, что маленькие дети, у которых организация коры идет наиболее активно, вообще, большую часть времени проводят во сне.
Идеи консолидации позволяют объяснить не только сон младенцев, но и консерватизм стариков. Под консерватизмом мы понимает отказ от новых идей и навыков при условии сохранения нормальной событийной памяти. В традиционных подходах это вызывает определенное противоречие. Казалось бы, способность запоминать новые факты должна обеспечивать способность обучаться новым принципам и осваивать новые навыки. Но все встает на место, если вспомнить, что есть организация нейронов-детекторов, формирующая набор понятий мозга, а есть событийная память, описывающаяся и хранящаяся на базе этих понятий. Ежедневная консолидация событийной памяти расчищает, по-видимому, больше половины доступного объема. Это значит, что независимо от возраста событийная память может быть крепкой, если этому не мешают органические нарушения. Но вот с нейронами-детекторами все обстоит сложнее. Похоже, что с возрастом количество нейронов, определившихся со своей специализацией, становится все больше, и для образования новых понятий остается все меньше и меньше возможностей. И тогда мы приходим к ситуации, что запомнить мы можем все, но только в тех понятиях, которые у нас существуют. А вот сформировать новые понятия мы уже можем с трудом или не можем вообще. Соответственно, все принципиально новое вызывает трудности восприятия и естественное отторжение, что и называется старческим консерватизмом.
Гиппокамп
Полное удаление гиппокампа делает невозможным формирование новых воспоминаний, что убедительно продемонстрировал случай с пациентом H.M. Нарушения в работе гиппокампа могут привести к синдрому Корсакова, который так же сводится к невозможности фиксировать текущие события, при сохранении старой памяти. Все это убеждает в том, что гиппокамп играет ключевую роль в механизме памяти.
Традиционные теории о роли гиппокампа строятся на аналогии мозга и компьютера. В таких рассуждениях гиппокампу отводится роль «оперативной памяти», то есть места, где накапливаются новые воспоминания. Затем же, предположительно во сне, эти воспоминания переносятся в области мозга, ответственные за хранение долговременной памяти. И хотя механизмы такого переноса непонятны, но, по крайней мере, это позволяет объяснить, почему нарушение работы гиппокампа блокирует формирование событийной памяти.
Наша модель принципиально отличается от моделей традиционных. У нас не происходит никакой буферизации памяти в гиппокампе и последующего ее копирования. Элементы, образующие воспоминания, сразу формируются там, где им уместно находиться. Гиппокамп же при этом просто создает единый идентификатор, который объединяет распределенные по пространству коры элементы памяти. Волновая модель работы мозга объясняет, как этот идентификатор распространяется по всей коре. Наличие такого идентификатора позволяет, выбрав объединенные им элементы, воспроизвести не абстрактную картину, а конкретный образ запомненного события.
В 2010 году были проделаны интересные опыты по протезированию гиппокампа крысы (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011). Подопытным животным с обеих сторон мозга в гиппокамп внедрялись микроэлектродные массивы, захватывающие области CA3 и CA1. После того, как крысы восстанавливались после операции, начинались серии тестов.
Крыс помещали в камеру с двумя выдвигающимися рычагами. Тест состоял из трех фаз. Сначала случайным образом выдвигался один из рычагов. Когда крыса нажимала этот рычаг, фиксировалась картина активности предварительно отобранных нейронов гиппокампа (стадия образца). Затем рычаг убирался, и наступала пауза от 1 до 30 секунд в одних сериях и до 60 в других. После чего выдвигались оба рычага. При нажатии на рычаг, противоположный исходному образцу, выдавалось подкрепление в виде капельки воды. При ошибке, то есть нажатии на тот же рычаг, что был на стадии образца, следовало наказание – выключение света на 5 секунд. В результате составлялся график зависимости успешности попыток от времени задержки (рисунок ниже).
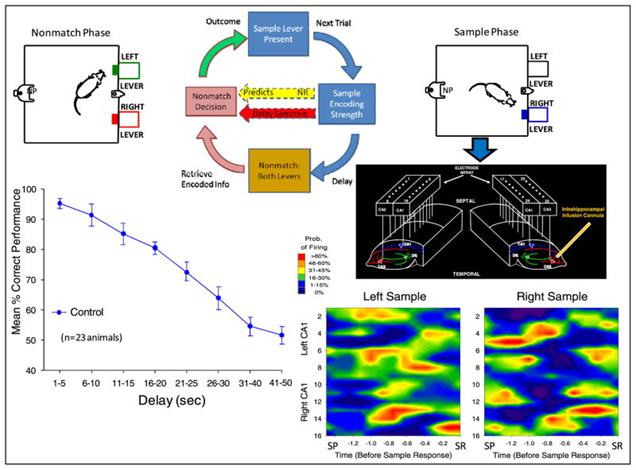
Контрольные измерения при естественной активности гиппокампа (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Исходя из записанных сигналов, была составлена предсказательная модель, которая прогнозировала, какой сигнал следует ждать на CA1 в зависимости от сигнала на CA3. В серии опытов сигналы прогноза были добавлены к существующим сигналам (рисунок ниже).

Добавление искусственных сигналов (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Получился интересный результат. Оказалось, что существенное влияние на результат оказывается только при корректировке сигнала в стадии предъявления образца, то есть при запоминании исходного образа. Картина же сигналов в остальные моменты была не принципиальна. Причем, чем сильнее был корректирующий сигнал, тем лучше был итоговый результат.
Авторы эксперимента, объясняя результат, исходили из представления, что гиппокамп занимается кодированием текущей информации, подготавливая ее для записи в память. Что картины активности, наблюдаемые в гиппокампе, несут информационное описание происходящих событий. Соответственно, усиление «правильного» сигнала способствует улучшению результатов запоминания.
В нашей модели объяснение полученных результатов несколько иное. Понятно, что идентификатор гиппокампа важен только в момент формирования памяти. После того как воспоминание создано гиппокамп не требуется для его воспроизведения. Усиление текущего идентификатора гиппокампа положительно сказывается на силе запоминания, но это не результат создания более четкой информационной картины, а просто выделение текущего воспоминания на фоне остальной памяти. Если у нормально работающего мозга таким образом подчеркивать все воспоминания, то итоговый результат окажется нулевой.
Самая интересная серия опытов была связана с подавлением собственной активности гиппокампа. Для этого крысам делались регулярные инъекции в область CA3 гиппокампа препарата MK801. MK801 блокирует нормальную работу синаптической передачи, использующую глутамат. Он проникает в ионные каналы NMDA рецепторов, чувствительных к глутамату и нарушает их работу.
Блокада участка CA3 ожидаемо резко ухудшила результаты. Но подача искусственных сигналов, восстанавливающих ожидаемую картину активности, позволила значительно улучшить количество правильных ответов (рисунок ниже).
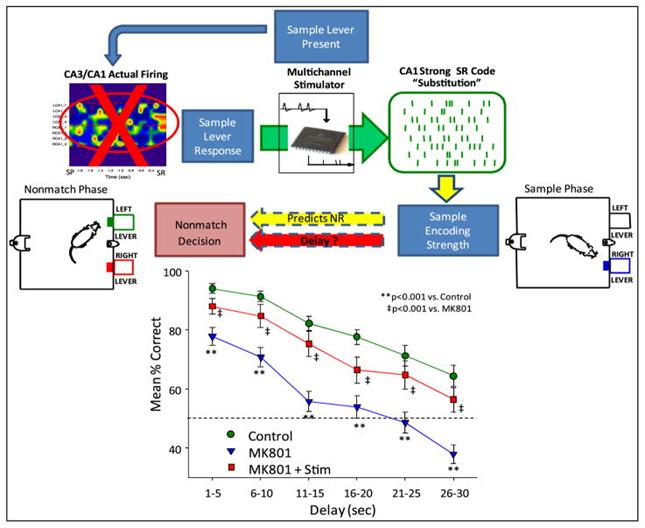
Замена естественного сигнала гиппокампа искусственным (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Я не склонен толковать этот результат как чудесное восстановление нейронного описания происходящего в гиппокампе с последующей записью этого описания в память. Тем более что в опыте использовались матрицы со всего 32 электродами, из которых задействовано было около половины. Скорее всего, происходило создание случайного идентификатора, который и делал возможным формирование памяти.
Но не все так просто с гиппокампом. Есть несколько фактов, которые достаточно сильно озадачивают при первом знакомстве с ними. Так в 1971 году Джон О’Киф открыл в гиппокампе клетки места (O’Keefe J., Dostrovsky J., 1971). Эти клетки реагируют подобно внутреннему навигатору. Если крысу поместить в длинный коридор, то по активности определенных клеток можно будет точно сказать, в каком его месте она находится. Причем реакция этих клеток не будет зависеть от того, как она попала в это место.
В 2005 году в гиппокампе были обнаружены нейроны, кодирующие положение в пространстве, образующие нечто наподобие координатной сетки (Hafting T., Fyhn M., Molden S., Moser M.B., Moser E.I., 2005).
В 2011 году оказалось, что в гиппокампе есть клетки, которые определенным образом кодируют временные интервалы. Их активность образует ритмические узоры, даже если ничего другого вокруг не происходит (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011).
Из всех этих фактов напрашивается вывод, что гиппокамп отвечает за кодирование нашего положения, как во времени, так и в пространстве. При этом гиппокамп подобен солдату, для которого вполне естественно «копать от забора и до обеда». Гиппокамп соединяет в себе и пространственную, и хронологическую навигацию, которые, кстати, имеют много общего. Так же, как наше путешествие в пространстве происходит от ориентира к ориентиру, так и путешествие во времени проходит от одной хронологической метки к другой.
Мы ранее утверждали, что гиппокамп формирует уникальные идентификаторы для воспоминаний. Как соотносится эта функция с тем, что гиппокамп отслеживает наши пространственно временные координаты? Конечно, можно считать, что гиппокамп несет двойную функцию, участвуя в двух независимых процессах. Но более логично предположить, что мы имеем дело с двумя сторонами одной медали.
Как мы идентифицируем информацию вообще? Используя Google, мы составляем запрос, состоящий набора слов, который определяет смысл нашего поиска. Получив список результатов, мы затем можем уточнить его, введя ограничения по дате или по географии.
Создавая библиотеку изображений, мы помечаем фотографии датой их создания и геотегами места, где они сделаны. Потом мы описываем, кто или что запечатлено на них.
Размещая файлы на компьютере, мы указываем время создания и путь к файлу. Названием файла мы кратко описываем его содержание.
Получается, что идентифицируя самые разные вещи, мы используем, с одной стороны, признаки, описывающие их смысл, а, с другой стороны, координаты в пространстве и времени, как универсальные признаки любых явлений. Такая идентификация впоследствии оказывается достаточно удобной и полезной не только как уникальная метка, но и как инструмент поиска. Можно предположить, что и природа пошла аналогичным путем.
То есть, создавая уникальный идентификатор события, гиппокампу имеет смысл не просто генерировать случайный код и распространять его по всей коре, а закладывать в этот код пространственно-временные признаки, как наиболее подходящие для всех случаев жизни. Такое пространственно-временное описание само по себе оказывается достаточно уникально, как сочетание разнообразных факторов. Достаточно добавить к нему некий небольшой случайный довесок и получившийся идентификатор, будет однозначно указывать на конкретное событие.
Такая идентификация значительно мощнее по своим возможностям, чем просто уникальный ключ для каждого воспоминания. Появляется возможность разнообразных дополнительных ассоциаций. Но самое главное, появляется возможность кодирования сложных описаний, учитывающих то, как развиваются события и во времени, и в пространстве. Но разговор об этом будет несколько позже.
Если допустить, что идентификатор гиппокампа действительно кодирует пространственно-временные координаты, то получается, что гиппокамп, во-первых, обладает способностью взаимодействовать с корой таким образом, чтобы иметь возможность обучаться узнавать похожие позиции в пространстве и времени. Во-вторых, чтобы распространять свои идентификаторы по всему пространству коры, гиппокамп должен формировать их из конечного числа уникальных фрагментов. Это проистекает из того, что кора должна быть предварительно обучена на распространение всего «алфавита», используемого гиппокампом. Новые идентификаторы должны формироваться не «с нуля», а как комбинации уже известных элементов. В принципе, в этих требованиях нет ничего сложного, это соответствует тому, как в описаниях на естественном языке все фразы строятся из конечного набора букв.
Ассоциативность памяти
Запомнить на активных нейронах-детекторах сопутствующую волновую активность, образующую уникальный идентификатор, – это только одна из составляющих памяти. Кроме того, надо уметь искать нужные воспоминания и обладать возможностью их воспроизведения.
Когда мы говорили о распространении нейронных волн, мы исходили из того, что нейрон хранит на внесинаптической мембране те волновые картины, участником которых он является. Встретив знакомую картину, нейрон своим спайком создает продолжение уникального узора. И тут важно, что нейрон не просто в состоянии узнать волновую картину, а то, что он сам – часть распространяющегося узора. Только будучи сам частью уникальной волны нейрон способен участвовать в ее распространении.
Чтобы не запутаться в последующих рассуждениях, еще раз повторим основные свойства волновой модели коры. Если отметить нейроны, относящиеся к одному волновому узору или – другими словами – идентификатору, то получится что-то вроде набора точек, изображенного на рисунке ниже.

Взяв любое место коры и активировав элементы идентификатора, мы получим волну, распространяющуюся от активного места, повторяющую характерный для идентификатора узор (рисунок ниже).

Проходя через каждое место коры, волна будет «высвечивать» фрагмент своего уникального узора. Так, стартовав из области 1, волна, дойдя до области 2, создаст там свой предопределенный идентификатором уникальный узор (рисунок ниже).

По уникальности узора можно в каждом месте коры определить, какие идентификаторы составляют волну.
Если мы на спокойной коре в области 2 воспроизведем уже знакомый нам узор, то он также создаст волну, которая, распространившись до области 1, создаст там все тот же паттерн, характерный именно для этого идентификатора.

Из всего этого следует, что для узнавания идентификатора достаточно в любом месте коры запомнить, какой узор именно в этом месте создает волна. Это можно запомнить либо на синапсах нейрона, либо на внесинаптической части мембраны. Запоминание на синапсах приводит к вызванной активности (пакету импульсов) при узнавании, запоминание на метаботропных рецептивных кластерах позволяет получать единичные спайки при появлении знакомой волны.
Сложнее, если нам надо воспроизвести идентификатор. Для этого нам надо активировать хотя бы единичными спайками группу близко расположенных нейронов, относящихся к требуемому идентификатору. Описывая обратную проекцию и вводя несколько медиаторов, мы как раз показывали механизм, реализующий нечто подобное.
Говоря об ассоциативности понятий, мы показали, что запрос, построенный на волнах идентификаторов, возвращает набор идентификаторов понятий, ассоциативно связанных с понятиями, содержащимися в запросе.
Чтобы показать механизм извлечения воспоминаний из ассоциативной событийной памяти нам надо показать, как волновой запрос, состоящий из понятий-признаков, может выдать уникальные гиппокамповские идентификаторы воспоминаний, подходящих под этот запрос. Если мы сможем получить такой набор идентификаторов и отобрать из них один, то, запустив этот идентификатор обратно по коре, мы получим активность нейронов-детекторов, содержащих этот идентификатор на внесинаптической мембране, а это будет равносильно восстановлению всей описательной картины требуемого воспоминания.
Напомню, про нашу упрощенность и схематичность описания. Далее я изложу возможный нейронный механизм ассоциативной памяти, не утверждая при этом, что мозг работает именно так.
Опять ненадолго обратимся к строению реальных нейронов. Тело нейрона – сома – имеет ограниченную площадь и не может обеспечить места для всех синаптических контактов. Большая часть синапсов приходится на разветвленную структуру, называемую дендритом или дендритным деревом (рисунок ниже). Количество синапсов, располагающихся на дендрите, в 10-20 раз превышает количество синапсов на соме.
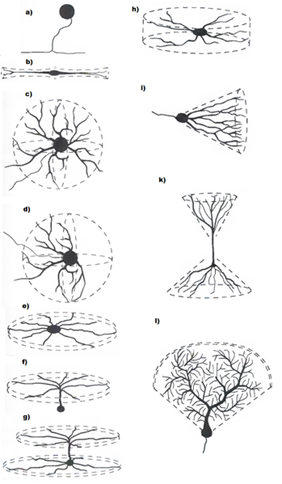
Формы дендритных деревьев (Greg Stuart, Nelson Spruston, Michael Häusser)
Было обнаружено, что нейрон ведет себя по-разному в зависимости от того, приходят ли сигналы на синапсы на одну или на разные дендритные ветки (Shepherd G.M., Brayton R.K., Miller J.P., Segev I., Rinzel J., Rall W., 1985). Одновременный приход импульсов на синапсы одной ветки вызывает значительно более сильный ответ нейрона, чем сигнал, распределенный по разным веткам.
На основе наблюдений такого рода родилась гипотеза о том, что дендритные ветки могут играть роль детекторов совпадений (Softky, 1994). Ее суть в том, что сигналам, рождающимся в удаленных ветках дендритного дерева, для генерации спайка нейрона необходимо, чтобы были активны сегменты дендритного дерева, лежащие по дороге сигнала к соме.
Такой эффект был показан для пирамидальных нейронов (Jarsky T., Alex Roxin A., Kath W.L., Spruston N., 2005), но можно полагать, что что-то подобное свойственно и нейронам других типов.
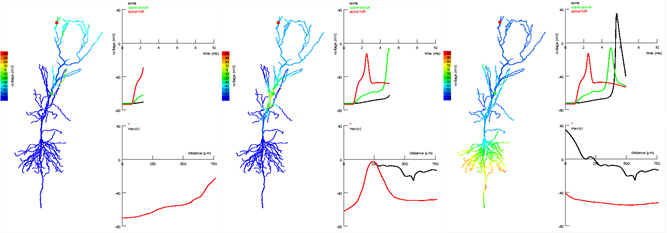
Фазы распространения сигнала в апикальных и наклонных сегментах пирамидального нейрона гиппокампа. Сигнал возникает в апикальном пучке (красная точка) и распространяется до коллатерали Шаффера (зеленая точка). Сигнал в более близком к соме месте дендрита не позволяет далекому сигналу угаснуть и способствует возникновению спайка (Jarsky T., Alex Roxin A., Kath W.L., Spruston N., 2005)
Распространение импульса по дендриту сопровождается его значительным затуханием. По идее, влияние удаленных (дистальных) синапсов должно быть значительно меньше, чем влияние близких (проксимальных). Однако были показаны механизмы, выравнивающие вклад таких синапсов, что вылилось в концепцию «демократии синапсов» (Clifton C. Rumsey, L. F. Abbott, 2006 ). Выравнивание вклада синапсов вдоль ветки дендрита позволяет рассматривать ветки как самостоятельные логические элементы, сигналы которых каким-то образом далее обрабатываются нейроном. Это означает, что теоретически, меняя конфигурацию дендрита и реакцию сомы, можно получить нейроны с различными логическими свойствами.
Например, в популярной концепции «иерархической темпоральной памяти» Джеффа Хокинса применяются нейроны, использующие автономные элементы, работающие в режиме «или» (рисунок ниже).
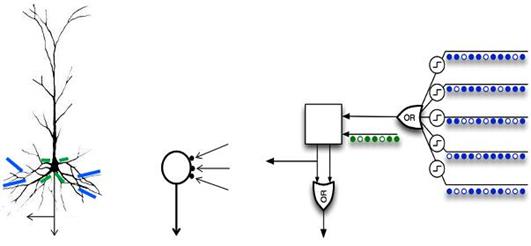
Модель нейрона с набором дендритных логических автономных элементов в сопоставлении с пирамидальным нейроном (Хокинс, 2011)
Вполне уместно предположить, что и реальный мозг оперирует нейронами с существенным разнообразием свойств.
Теперь перейдем, собственно, к описанию ассоциативной событийной памяти. Предположим, что у нас есть нейроны двух типов, образующих плоскую кору. Рецептивные поля этих нейронов охватывают некую локальную область своего окружения, куда попадают нейроны обоих типов. Условно разнесем их на два слоя, помня при этом о перекрестном распределении связей.
Зададим нейроны первого типа такими, чтобы они распространяли волну информационного идентификатора. Нейроны же второго типа заставим распространять исключительно волны идентификаторов гиппокампа (рисунок ниже).
Распространение двух независимых волн на нейронах с разным типом медиатора
Для этого разведем их аксоны и внесинаптические рецепторы по используемым медиаторам (таблица ниже). Обратите внимание, что для задания волнового распространения синаптические рецепторы нам не важны.
|
Синапсы |
Вне синапсов |
Аксон |
|
| Тип 1 |
A |
А |
|
| Тип 2 |
B |
B |
Нейромедиаторные характеристики нейронов. Чувствительность рецепторов и выброс аксона
При таком задании медиаторов и рецепторов волны на двух слоях никак не будут влиять друг на друга.
Теперь усложним нейроны второго типа, сделаем их дендритные деревья состоящими из двух типов веток, отличающихся внесинаптическими медиаторами. При моделировании однородные ветки можно объединить и в результате оставить два дендритных сегмента, работающих по принципу «или» (таблица ниже).
|
Сегмент |
Синапсы |
Вне синапсов |
Аксон |
|
| Тип 1 |
A |
А |
||
| Тип 2 |
1 |
B |
B |
|
|
2 |
A |
Нейромедиаторные характеристики нейронов. Чувствительность рецепторов и выброс аксона. Нейроны второго типа имеют два дендритных сегмента с рецепторами, чувствительными к медиаторам разного типа
Предположим, что пока шло обучение нейронов тому, как распространять волны идентификаторов, у нейронов второго типа был заблокирован второй сегмент, чувствительный к «чужому» медиатору. Вся информация, необходимая для формирования волн, у нейронов второго типа будет откладываться на внесинаптической памяти первого сегмента.
Если теперь мы включим вторые сегменты, то на них начнется запоминание волновых картин, распространяющихся по нейронам первого типа. Будем полагать, что внесинаптическое запоминание происходит в момент, когда нейрон генерирует спайк. При этом фиксируется не моментальная картина активности на синапсах, а накопленная за весь такт распространения волны. Накопление позволит нам запомнить фрагмент волны первого уровня, даже если фазы волн двух уровней в месте запоминания не совпадают. То есть, когда нейрон второго уровня выдаст спайк, он зафиксирует картину информационной активности, даже если она была какое-то время назад.
Предположим, что мы провели один такт такого запоминания. Все нейроны второго уровня, распространяющие волну идентификатора гиппокампа, активные в этом такте, запомнят на своих вторых сегментах фрагменты информационной волны, прошедшей по нейронам первого типа. Это значит, что если мы когда-нибудь повторим такую же волну описания, то все нейроны второго уровня, которые узнают эту картину, дадут спайк. Общая картина этих спайков воспроизведет тот самый идентификатор гиппокампа, который был на момент запоминания этого информационного образа.
Так можно запоминать различные описательные картины. При повторении соответствующих описаний нейроны второго уровня будут выдавать суммарное сочетание всех идентификаторов гиппокампа, попадающих под текущее описание.
Видно, что такое запоминание крайне расточительно. Мы запоминаем каждый образ на всех активных нейронах второго типа. Хотя для воспроизведения идентификатора нам нужен всего лишь малый локальный участок, способный запустить волну. Чтобы исправить ситуацию, будем запоминать информационную волну не на всех активных нейронах второго типа, а только на тех, которые попадают в области с высокой вызванной активностью нейронов первого типа. То есть вспомним, что информация дуалистична. Она одновременно и идентификаторная волна, и активность паттернов вызванной активности. Волна даст нам узор для запоминания, а паттерны укажут места для этого.
Самый простой способ выбора места – это воспользоваться тем же полем активности, которое мы использовали для пространственной самоорганизации паттернов. В такой конфигурации запоминание можно представить так. В местах коры, где информационная картина создает паттерны вызванной активности, формируется поле активности. Все активные нейроны второго типа, распространяющие волну гиппокампа, для которых поле активности выше определенного порога, фиксируют на внесинаптической памяти картины, описываемые информационной волной.
Подав информационную волну-запрос, мы получим активность нейронов второго типа, узнавших этот запрос. Локальные группы таких нейронов будут излучать идентификаторы гиппокампа, относящиеся ко всем воспоминаниям, ассоциативно связанным с этим запросом.
Мы показали, как могут фиксироваться и восстанавливаться уникальные идентификаторы воспоминаний. Ранее мы показали, что если на нейронах-детекторах хранить память о таких уникальных идентификаторах, то можно восстанавливать образы, соответствующие запомненным моментам. В таблице ниже приведена возможная конфигурация медиаторов для такой памяти.
|
Сегмент |
Синапсы |
Вне синапсов |
Аксон |
|
| Тип 1 |
1 |
A |
А |
|
|
2 |
B |
|||
| Тип 2 |
1 |
B |
B |
|
|
2 |
A |
Нейромедиаторные характеристики нейронов. Чувствительность рецепторов и выброс аксона. Двусвязная конфигурация
Задание области запоминания через поле активности – достаточно грубый инструмент. Есть вполне очевидные пути по совершенствованию конструкции. Например, можно через синаптическое обучение нейронов второго типа создать из них области, соответствующие понятиям, находящимся в этом месте коры. Используя концепцию дендритного детектора совпадений, можно сделать так, что активность понятия будет обязательным условием, как для записи, так и для обратной генерации идентификаторов воспоминаний, связанных с этим понятием.
Вообще же богатство инструментария, которое предоставляет волновая модель коры с многосекционными нейронами, позволяет реализовывать достаточно сложные и интересные конструкции. Ограничимся пока уже приведенными самыми простыми схемами, чтобы окончательно не запутать повествование.
Следы памяти
Энграммой называют те изменения, что происходят с мозгом в момент запоминания. Другими словами, энграмма – это след памяти. Вполне естественно, что понимание природы энграмм воспринимается всеми исследователями как ключевая задача в изучении природы мышления.
В чем сложность этой задачи? Если взять обычную книгу или внешний компьютерный накопитель, то и то и другое можно назвать памятью. И то и другое хранит информацию. Но мало хранить. Чтобы информация стала полезной, надо уметь ее считывать и знать, как ей оперировать. И тут оказывается, что сама форма хранения информации тесно связана с принципами ее обработки. Одно во многом определяет другое.
Человеческая память – это не просто способность хранить большой набор разнообразных образов, но еще и инструмент, позволяющий быстро найти и воспроизвести уместное воспоминание. При этом кроме ассоциативного доступа к произвольным фрагментам нашей памяти мы умеем связывать воспоминания в хронологические цепочки, воспроизводя не единичный образ, а последовательность событий.
Уайлдер Грейвс Пенфилд получил заслуженное признание за свой вклад в изучение функций коры. Занимаясь лечением эпилепсий, он разработал методику операций на открытом мозге, по ходу которых использовалась электростимуляция, позволяющая уточнить эпилептический очаг. Возбуждая электродом различные участки мозга, Пенфилд фиксировал реакцию находящихся в сознании пациентов. Это позволило получить подробное представление о функциональной организации коры головного мозга (Пенфилд, 1950). Стимулирование некоторых зон, в основном височных долей, вызывало у пациентов яркие воспоминания, в которых прошедшие события всплывали в мельчайших подробностях. Причем повторное стимулирование одних и тех же мест вызывало одни и те же воспоминания.
Выявленная Панфилом четкая локализация в коре многих функций настроила на поиски таких же четко локализованных следов памяти. Кроме того, появление компьютеров и соответственно представления о том, как организованы физические носители компьютерной информации, стимулировали поиск чего-то похожего в структурах мозга.
В 1969 году Джерри Летвин сказал: «Если мозг человека состоит из специализированных нейронов, и они кодируют уникальные свойства различных объектов, то, в принципе, где-то в мозге должен быть нейрон, с помощью которого мы узнаем и помним свою бабушку». Формулировка «нейрон бабушки» закрепилась и часто всплывает, когда заходит разговор об устройстве памяти. Более того, нашлись и прямые экспериментальные подтверждения. Были обнаружены нейроны, реагирующие на определенные образы, например, четко узнающие конкретного человека или конкретное явление. Правда, при более детальных исследованиях оказалось, что обнаруженные «специализированные» нейроны реагируют не только на что-то одно, а на группы в некотором смысле близких образов. Так, выяснилось, что нейрон, реагировавший на Дженнифер Энистон, так же реагировал и на Лизу Кудроу, снимавшуюся вместе с Энистон в телесериале «Друзья», а нейрон, узнававший Люка Скайуокера, узнавал и мастера Йоду (Р. К. Квирога, К. Кох, И. Фрид, 2013).
Еще в первой половине двадцатого века Карл Лэшли поставил очень интересные опыты по локализации памяти. Сначала он обучал крыс находить выход в лабиринте, а затем удалял им различные части мозга и снова запускал в лабиринт. Так он пытался найти ту часть мозга, которая отвечает за память о полученном навыке. Но оказалось, что память в той или иной мере сохранялась всегда, несмотря на временами значительные нарушения моторики. Эти опыты вдохновили Карла Прибрама сформулировать ставшую широко известной и популярной теорию голографической памяти (Прибрам, 1971).
Принципы голографии, как и сам термин, придумал в 1947 году Денеш Габор, который получил за это Нобелевскую премию по физике 1971 года. Суть голографии в следующем. Если иметь источник света со стабильной частотой, то разделив его посредством полупрозрачного зеркала на два, мы получим два когерентных световых потока. Один поток можно направить на объект, а второй на фотографическую пластину.
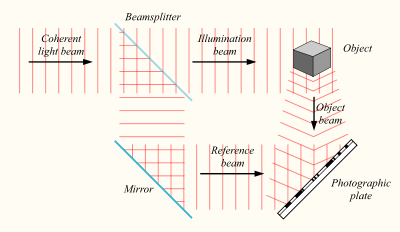
Создание голограммы
В результате, когда отраженный от объекта свет достигнет фотографической пластины, он создаст с освещающим пластину потоком интерференционную картину.
Интерференционная картина, отпечатавшись на фотопластинке, сохранит информацию не только об амплитудных, но и о фазовых характеристиках светового поля, отраженного объектом. Теперь, если осветить ранее экспонированную пластину, то восстановится исходный световой поток, и мы увидим запомненный объект во всем его объеме.
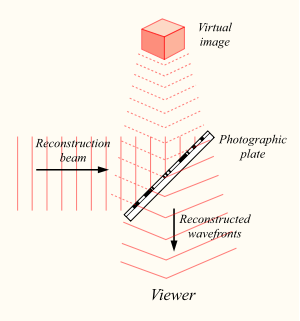
Воспроизведение голограммы
Голограмма имеет несколько удивительных свойств. Во-первых, световой поток сохраняет объем, то есть, разглядывая фантомный объект под разными углами, можно увидеть его с разных сторон. Во-вторых, каждый участок голограммы содержит в себе информацию обо всем световом поле. Так, если мы разрежем голограмму пополам, то сначала мы увидим только половину объекта. Но если мы наклоним голову, то за краем оставшейся голограммы мы сможем разглядеть вторую «обрезанную» часть. Да, чем меньше фрагмент голограммы, тем ниже ее разрешающая способность. Но даже через маленький участок можно, как через замочную скважину, рассмотреть все изображение. Интересно, что если на голограмме будет лупа, то через нее можно будет рассматривать с увеличением другие запечатленные там объекты.
Применительно к памяти Прибрам сформулировал: «Сущность голографической концепции состоит в том, что образы восстанавливаются, когда их представительства в виде систем с распределенной информацией соответствующим образом приводятся в активное состояние» (Прибрам, 1971).
Упоминание о голографических свойствах памяти можно встретить в двух контекстах. С одной стороны, называя память голографической, подчеркивают ее распределенный характер и способность восстанавливать образы, используя только часть нейронов, подобно тому, как это происходит с фрагментами голограммы. С другой стороны, предполагают, что память, обладающая похожими на голограмму свойствами, опирается на те же физические принципы. Последнее означает, что поскольку в основе голографии лежит фиксация интерференционной картины световых потоков, то память, видимо, каким-то образом использует интерференционную картину, возникающую в результате импульсного кодирования информации. Ритмы мозга хорошо известны, а там, где колебания там и волны, а, следовательно, неизбежна и их интерференция. А значит, физическая аналогия выглядит вполне уместной и привлекательной.
Но интерференция штука тонкая, небольшие изменения частоты или фазы сигналов должны полностью менять ее картину. Однако мозг успешно работает и при существенной вариации его ритмов. Кроме того, попытки затруднить распространение электрической активности рассечением его участков и помещением слюды в места разрезов, накладыванием полосок золотой фольги для создания замыкания, созданием эпилептических очагов посредством инъекций алюминиевой пасты не нарушают слишком уж паталогически деятельность мозга (Прибрам, 1971).
Говоря о памяти, невозможно обойти стороной известные факты о связи памяти и гиппокампа. В 1953 году пациенту, которого принято называть H.M. (Henry Molaison), хирург удалил гиппокамп (W. Scoviille, B. Milner, 1957). Это была рискованная попытка вылечить сильную эпилепсию. Было известно, что удаление гиппокампа одного из полушарий действительно помогает при этой болезни. Учитывая исключительную силу эпилепсии у H.M., врач удалил гиппокамп с обеих сторон. В результате у H.M. полностью исчезла способность что-либо запоминать. Он помнил, что было с ним до операции, но все новое вылетало у него из головы, как только переключалось его внимание.

Henry Molaison
H.M. долго исследовался. По ходу этих исследований было проведено бесчисленное количество разных опытов. Один из них оказался особенно интересен. Пациенту предложили обвести пятиконечную звезду, глядя на нее в зеркало. Это не очень простое задание, вызывающее затруднение при отсутствии должного навыка. Задание давалось H.M. неоднократно и каждый раз он воспринимал его, как увиденное впервые. Но интересно, что с каждым разом выполнение задания давалось ему все легче и легче. Он сам при повторных опытах отмечал, что ожидал, что это окажется значительно сложнее.
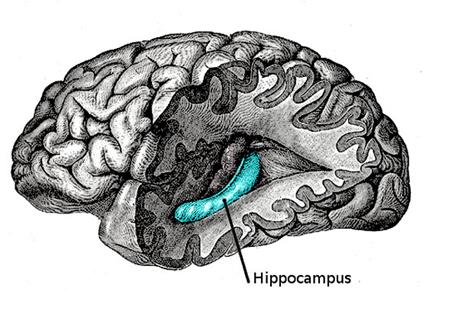
Гиппокамп одного из полушарий
Кроме того, оказалось, что определенная память на события все-таки была присуща H.M. Так, например, он знал об убийстве Кеннеди, хотя оно и произошло уже после удаления у него гиппокампа.
Из этих фактов был сделан вывод, что существует как минимум два различных типа памяти. Один тип отвечает за фиксацию конкретных воспоминаний, а другой ответственен за получение некого обобщенного опыта, что выражается в знании распространенных фактов или приобретении определенных навыков.
Случай H.M. достаточно уникален. В других ситуациях, связанных с удалением гиппокампа, где не было такого полного двустороннего повреждения, как у H.M., нарушения памяти либо были не столь ярко выражены, либо отсутствовали вовсе (W. Scoviille, B. Milner, 1957).
Попробуем теперь сопоставить все описанное с нашей моделью. Мы показали, что устойчивые повторяющиеся явления формируют паттерны нейронов детекторов. Эти паттерны способны узнавать характерные для них сочетание признаков, и добавлять в волновую картину новые идентификаторы. Мы показали, как может происходить обратное воспроизведение признаков по идентификатору понятия. Это можно сопоставить с памятью об обобщенном опыте.
Но такая обобщенная память не позволяет воссоздать конкретные события. Если одно и то же явление повторяется в разных ситуациях, мы в нашей нейронной сети просто получаем ассоциативные связи между понятием, соответствующим явлению, и понятиями, описывающими эти обстоятельства. Используя эту ассоциативность, можно создать абстрактное описание, состоящее из понятий, встречающихся совместно. Задача же событийной памяти – это не воспроизвести некую абстрактную картину, а воссоздать именно запомненную ранее ситуацию, описывающую конкретное событие со всеми только ему характерными уникальными особенностями.
Сложность собственно в том, что в нашей модели нигде нет такого места, где бы локализовалось полное и исчерпывающее описание происходящего. Полное описание складывается из множества описаний, активных на отдельных зонах коры. Каждая из зон имеет волновое описание в понятиях, которые свойственны именно этому участку мозга. И если мы даже как-то запомним, что происходит на каждой из зон в отдельности, эти описания будет необходимо еще увязать между собой так, чтобы возник целостный образ.
Аналогичная ситуация возникает, когда мы имеем топографическую проекцию и нейроны с локальными рецептивными полями. Предположим, что у нас есть нейронная сеть, состоящая из двух плоских слоев (рисунок ниже). Допустим, что состояние нейронов первого слоя образует определенную картинку. Эта картинка через проекционные волокна передается на второй слой. Нейроны второго слоя имеют синаптические связи с теми волокнами, которые попадают в границы их рецептивных полей. Таким образом, каждый из нейронов второго слоя видит только маленький фрагмент исходной картинки первого слоя.
Топографическая проекция образа на локальные рецептивные поля
Есть очевидный способ, как запомнить на втором слое подаваемую картинку. Надо выбрать такой набор нейронов, чтобы их рецептивные поля полностью покрывали проецируемый образ. Запомнить на каждом из нейронов свой фрагмент изображения. А чтобы воспоминание приобрело связанность – пометить все эти нейроны общим маркером, указывающим на их принадлежность к одному набору.
Такое запоминание очень просто, но крайне расточительно по количеству задействованных нейронов. Каждая новая картинка будет требовать новый распределенный набор элементов памяти.
Получить экономию можно, если окажется, что у разных изображений повторяются какие-либо общие фрагменты, тогда можно не заставлять новый нейрон запоминать такой фрагмент, а воспользоваться уже существующим нейроном, просто добавив ему еще один маркер, теперь уже от новой картинки.
Таким образом, мы приходим к основной идее распределенного запоминания. Опишем ее сначала для картинки и топографической проекции.
Будем подавать на первую зону различные изображения и проецировать их на вторую зону. Если мы сделаем рецептивные поля нейронов достаточно малыми, то количество уникальных картинок в каждой локальной области окажется не столь велико. Мы можем так подобрать размеры рецептивного поля, чтобы в область, размеры которой будут приблизительно совпадать с размером рецептивного поля нейронов, поместились все уникальные варианты локальных изображений.
Создадим пространственные области, содержащие нейроны-детекторы. Сделаем так, чтобы каждая область содержала детекторы всех возможных уникальных образов и чтобы такие области покрывали все пространство второй зоны. Для этого мы можем воспользоваться описанными ранее принципами выделения наборов факторов.
Задача детекторов – сравнивать подаваемые на их рецептивные поля образы с образами, характерными для них. Для такого сравнения образов можно использовать свертку по рецептивному полю R:
\[y=\sum_{i\in R}x_{i}\omega _{i}\]
Отклик нейрона будет тем выше, чем больше новый образ покрывает образ запомненный. Если нас интересует не степень покрытия, а уровень совпадения образов, то можно воспользоваться корреляцией образов, которая есть не что иное, как нормированная свертка:
\[y=\frac{\sum_{i}x_{i}\omega _{i}}{\sqrt{\sum_{i}x_{i}^2\sum_{i}\omega _{i}^2}}\]
Кстати, это же значение является косинусом угла, образованного вектором образа и вектором весов:
\[y=\frac{x^T\omega }{\left \| x \right \|\left \| \omega \right \|}\]
В результате, в каждой локальной группе детекторов при подаче новой картинки будут срабатывать нейроны-детекторы, наиболее точно описывающие свой локальный фрагмент.
Теперь сделаем следующее: будем генерировать для каждого нового образа свою уникальную метку-идентификатор и помечать ею активные нейроны-детекторы. Мы получим, что каждая подача образа сопровождается появлением на второй зоне коры картины активности, которая есть описание этого образа через доступные второй зоне признаки. Создание уникального идентификатора и маркировка им активных нейронов детекторов – это и есть запоминание конкретного события.
Если мы выберем один из маркеров, найдем нейроны-детекторы, его содержащие, и восстановим характерные для них локальные образы, то мы получим восстановление исходного изображения.
Чтобы запоминать и воспроизводить множество разных образов, нейроны-детекторы должны иметь неизменные синаптические веса и обладать возможностью хранить столько маркеров, сколько воспоминаний им предстоит запомнить.
Покажем работу распределенного запоминания на простом примере. Допустим, что мы генерируем на верхней зоне контурные изображения разнообразных геометрических фигур (рисунок ниже).
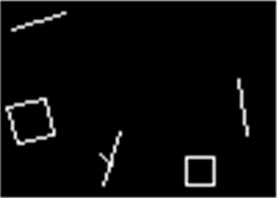
Подаваемое изображение
Будем обучать нижнюю зону на выделение различных факторов методом декорреляции. Основные образы, которые будут появляться в каждом малом рецептивном поле – это линии под разными углами. Будут и другие образы, например, пересечения и углы, свойственные геометрическим фигурам. Но линии будут доминировать, то есть встречаться чаще. Это значит, что они и выделятся в первую очередь в виде факторов. Реальный результат такого обучения показан на рисунке ниже.

Фрагмент поля факторов, выделенных из контурных изображений
Видно, что выделяется много вертикальных и горизонтальных линий, отличающихся своим положением на рецептивном поле. Это не удивительно, так как даже небольшое смещение создает новый фактор, не имеющий пересечений со своими параллельными «близнецами». Предположим, что мы каким-то образом усложнили нашу сеть так, что расположенные рядом параллельные «близнецы» слились в один фактор. Далее допустим, что в малых областях выделились факторы, как показано на рисунке ниже, с определенной дискретностью описывающие все возможные направления.
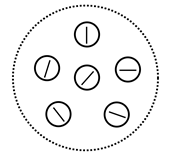
Факторы в малой области, соответствующие различным направлениям с дискретностью в один час
Тогда результат обучения всей зоны коры можно условно изобразить следующим образом:

Условный результат обучения зоны коры. Для наглядности нейроны размещены не на регулярной сетке
Теперь подадим на обученную зону коры изображение квадрата. Нейроны, которые увидят на своем рецептивном поле характерный для них стимул, активируются (рисунок ниже).
Реакция обученной зоны коры на образ квадрата
Теперь сгенерируем случайное уникальное число – идентификатор воспоминания. Для простоты не будем пока использовать наши волновые сети, ограничимся предположением, что каждый нейрон может хранить кроме синаптических весов еще и набор идентификаторов, то есть некий большой массив неупорядоченных чисел. Заставим все активные нейроны запомнить в своих наборах только что сгенерированный идентификатор. Собственно, этим действием мы и зафиксируем память об увиденном квадрате.
Подавая новые изображения, будем для каждого из них генерировать свой уникальный идентификатор и добавлять его к нейронам, отреагировавшим на текущий образ. Теперь, чтобы что-то вспомнить, достаточно будет взять соответствующий идентификатор, активировать все нейроны, которые его содержат, а затем восстановить картины образов, характерных для этих нейронов. Естественно, чем богаче и четче будет система описания, тем точнее восстановленный образ будет совпадать с исходным. Но даже на очень грубых моделях, например, на сети, приведенной выше, можно получать вполне правдоподобные результаты восстановления (рисунок ниже).
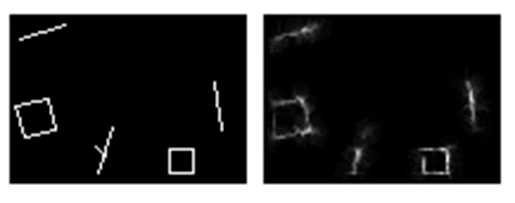
Исходное изображение и воспоминание, восстановленное по факторам «грубой» модели
Теперь можно сформулировать наши предположения относительно того, как устроена событийная память реального мозга.
Обучение различных зон коры ведет к формированию паттернов нейронов-детекторов, способных реагировать на образы, характерные для этих зон. В основе этого обучения лежит синаптическая пластичность. Это обучение не фиксирует конкретные события, а только выделяет обобщенные понятия. Когда возникает «нейрон бабушки», по его синаптическим весам нельзя судить о воспоминаниях, как-либо связанных с бабушкой. Веса синапсов описывают не конкретные события, а картины признаков, характерных для узнавания бабушки.
Картина описания, происходящего на каждой из зон коры, дуалистична. Это одновременно и картина вызванной активности нейронов-детекторов, и волновой идентификатор, сформированный как собственными для коры излучающими паттернами, так и волнами, пришедшими по системе проекций.
Роль гиппокампа – создавать уникальный идентификатор для каждого воспоминания и добавлять его в общую волновую картину. В результате этого на каждой зоне коры в волновом идентификаторе кроме перечисления признаков, описывающих текущее событие, появится еще и уникальная добавка от гиппокампа, которая позволит различить волновые описания похожих между собой событий.
Нейроны-детекторы, находящиеся в состоянии вызванной активности, фиксируют на своих метаботропных кластерах текущий волновой идентификатор. Кстати, мы уже наблюдали нечто похожее, когда описывали систему обобщенной ассоциативности понятий. Только теперь в идентификатор добавилась уникальная составляющая от гиппокампа. Этим действием мы создадим энграмму, позволяющую найти все нейроны-детекторы, относящиеся к одному воспоминанию.
Надо отметить, что эта конструкция одинаково работает независимо от того, как на зону коры поступает информация. И для топографической и для волновой проекции принципы запоминания остаются неизменны.
Такая конструкция памяти обладает всеми требуемыми от памяти голографическими свойствами. Нейроны-детекторы любого детекторного паттерна сохраняют информацию обо всей волновой картине, которая была на коре на момент запоминания, что в точности соответствует тому, как фрагмент голограммы хранит информацию о волновой картине окружающего пространства.
Фиксация энграммы получается распределенной по всем участкам мозга, проявившим активность в узнавании происходящего. Это значит, что воспоминание не привязано к какому-то одному или даже нескольким нейронам и не имеет конкретной локализации. Удаление любого участка коры, как это было в опытах Лэшли, не уничтожает всей энграммы, а только обедняет ее на описание в тех понятиях, которые оказались удалены.
Становится понятна природа нейронов, реагирующих на Дженнифер Энистон или мастера Йоду. Это не нейроны воспоминания – это лишь нейроны-детекторы, связанные с понятиями, имеющими отношение к соответствующим фильмам.
Можно объяснить природу ярких видений, возникающих при стимуляции мозга электродом. Электрод возбуждает случайный паттерн нейронов, с которыми он соприкасается. Если оказывается, что этот паттерн похож на фрагмент волны какого-либо известного коре идентификатора, то он запускает соответствующую волну, которая выстраивает всю остальную информационную картину мозга. Повторный электрический импульс в уже введенный электрод вызывает то же самое видение, так как создает тот же паттерн активности. Но при этом характер видения оказывается никак не связан с местом, куда попал электрод. Активируются не понятия, действительно имеющие локализацию, а волна, которая, в принципе, могла бы возникнуть в любом другом месте. Просто в месте введения электрода узор этой волны совпал с формой иглы. Для волновой модели коры все это вполне естественно, но вызывает озадаченность у тех, кто ищет «нейроны бабушки».
Наша концепция памяти хорошо объясняет особенности пациента H.M. Так как гиппокамп необходим для создания уникального идентификатора, то не удивительно, что его отсутствие сделало невозможным создание новых воспоминаний и при этом не нарушились воспоминания, уже имеющиеся. Там, где идентификатор уже был присвоен, гиппокамп больше не нужен для последующих информационных процедур. Так как формирование нейронов-детекторов и детекторных паттернов не завязано на гиппокамп, то объясняется и сохранение способности к процедурному обучению и формированию обобщенной памяти.
Еще раз обращу внимание на то, что при описании событийной памяти мы не использовали пластичность синапсов как инструмент запоминания. Пластичность синапсов у нас – это механизм формирования паттернов нейронов-детекторов. То есть, следы конкретных событий невозможно найти непосредственно на синапсах нейронов, хотя образы, описываемые синаптическими весами, и напоминают всегда что-то из встречавшегося ранее опыта. Мы пришли к необходимости разделения механизмов обучения и событийной памяти. Исходя их этого, в нашей модели получилось два типа энграмм. Один тип – это модификация синаптических весов, позволяющая выделиться признакам, на базе которых строятся все последующие описания. Второй тип – образование внесинаптических метаботропных рецептивных кластеров, объединяющих множество нейронов, участвующих в описании конкретного события. Причем второй тип энграмм невозможен без первого. Это означает, что для формирования воспоминаний о каких-либо событиях во всей полноте, должны существовать факторы, позволяющие составить такое описание.
Информация описывается мозгом иерархически, выделяя от уровня к уровню все более абстрактные признаки. Когда мы говорим о воспоминании относительно какого-либо события, мы, как правило, не имеем в виду фотографическую память нижнего уровня, разговор идет о фиксации более абстрактного описания, которое в процессе восстановления может привести к реконструкции исходной фотографической картины. Но чтобы такое описание было возможно, необходимо, чтобы были сформированы соответствующие факторы. Похоже, что именно по этой причине у нас нет ранних детских воспоминаний. В том возрасте, к которому относится провал нашей памяти, у нас просто еще нет тех понятий, которые необходимы для полноценного описания событий.
Динамические нейронные сети. Ассоциативность
Наиболее просты для понимания и моделирования нейронные сети, в которых информация последовательно распространяется от слоя к слою. Подав сигнал на вход, можно так же последовательно рассчитать состояние каждого из слоев. Эти состояния можно трактовать как набор описаний входного сигнала. Пока не изменится входной сигнал, останется неизменным и его описание.
Более сложная ситуация возникает, если ввести в нейронную сеть обратные связи. Чтобы рассчитать состояние такой сети, уже недостаточно одного прохода. Как только мы изменим состояние сети в соответствии с входным сигналом, обратные связи изменят входную картину, что потребует нового пересчета состояния всей сети, и так далее.
Идеология рекуррентной сети зависит от того, как соотносится задержка обратной связи и интервал смены образов. Если задержка много меньше интервала смены, то нас, скорее всего, интересуют только конечные равновесные состояния, и промежуточные итерации стоит воспринимать, как исключительно расчетную процедуру. Если же они сопоставимы, то на первый план выходит именно динамика сети.
Ф. Розенблатт описал многослойные персептроны с перекрестными связями и показал, что они могут быть использованы для моделирования избирательного внимания и воспроизведения последовательности реакций (Rosenblatt, Principles of Neurodynamic: Perceptrons and the Theory of Brain Mechanisms, 1962).
В 1982 году Джон Хопфилд предложил конструкцию нейронной сети с обратными связями такую, что ее состояние может быть определено, исходя из расчета минимума функционала энергии (Hopfield, 1982). Оказалось, что сети Хопфилда обладают замечательным свойством – они могут использоваться как память, адресуемая по содержанию.
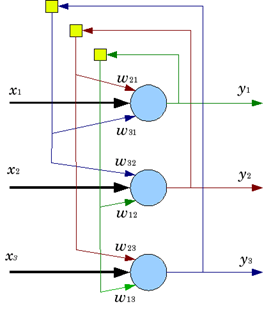
Сеть Хопфилда
Сигналы в сети Хопфилда принимают спиновые значения {-1,1}. Обучение сети – это запоминание нескольких образов \(X_{i}\). Запоминание происходит за счет настройки весов так, чтобы:
\[X_{i}=WX_{i}\]
Что приводит к алгоритму, который простым неитерационным расчетом позволяет определить параметры сети:
\[\omega _{ji}=\frac{1}{N}\sum_{k=1,,m}x_{ik}x_{jk}\]
Где \(\omega\)– матрица, составленная из весов нейронов, а \(m\) – количество запоминаемых образов. Диагональ матрицы \(\omega\) полагается нулевой, что означает отсутствие влияния нейронов на самих себя. Веса, заданные таким образом, определяют устойчивые состояния сети, соответствующие запомненным векторам.
Функция активации нейронов имеет вид:
\[a_{i}=\sum_{j}\omega _{ji}y_{i}\cdot \rightarrow y_{i}=\left\{\begin{matrix}1, & a_{i}\geq 0 \\ -1, &a_{i}< 0 \end{matrix}\right.\]
Входной образ присваивается сети, как начальное приближение. Затем начинается итерационная процедура, которая, если повезет, сходится к устойчивому состоянию. Таким устойчивым состоянием, скорее всего, будет один из запомненных образов. Причем это будет образ, наиболее похожий на входной сигнал. Другими словами, образ ассоциативно с ним связанный.
Можно ввести понятие энергии сети, как:
\[E=\frac{1}{2}\sum_{i,j=1}^{N}\omega _{ij}x_{i}x_{j}\]
Где \(N\) – количество используемых нейронов. Каждому запомненному образу в таком случае будет соответствовать локальный энергетический минимум.
Хотя сети Хопфилда и очень просты, они иллюстрирует три фундаментальных свойства мозга. Первое – это существование мозга в динамике, когда любые внешние или внутренние возмущения заставляют покинуть состояние текущего локального минимума энергии и перейти к динамическому поиску нового. Второе – это способность прийти к квазиустойчивому состоянию, определяемому предыдущей памятью. Третье – ассоциативность переходов, когда в смене описательных состояний постоянно прослеживается определенная обобщенная близость.
Как и сети Хопфилда, наша модель изначально динамична. Она не может существовать в статичном состоянии. Любая волна существует только в движении. Вызванная активность нейронов запускает волны идентификаторов. Эти волны вызывают активность паттернов, узнавших знакомые волновые сочетания. Паттерны запускают новые волны. И так без остановки.
Но динамика наших сетей принципиально отличается от динамики сетей Хопфилда. В традиционных сетях динамика сводится к итерационной процедуре схождения состояния сети к устойчивому состоянию. У нас же динамика сродни тем процессам, что протекают в алгоритмических компьютерах.
Волновое распространение информации – это механизм передачи данных. Динамика этой передачи – это не динамика итерационного схождения, а динамика пространственного распространения идентификационной волны. Каждый цикл распространения информационной волны меняет картину вызванной активности нейронов. Переход коры в новое состояние означает смену текущего описания и соответственно смену картины нашей мысли. Динамика этих изменений опять же не является некой итерационной процедурой, а формирует последовательность образов нашего восприятия и мышления.
Наша волновая модель значительно сложнее, чем простая динамическая сеть. Позже мы опишем достаточно много небанальных механизмов, регулирующих ее работу. Не удивительно, что и представление об ассоциативности в нашем случае оказывается сложнее, чем ассоциативность в сетях Хопфилда, и скорее напоминает концепции, свойственные компьютерным моделям представления данных.
В программировании под ассоциативностью понимается способность системы искать данные, похожие на образец. Например, если мы обращаемся к базе данных, указывая критерии запроса, то, по сути, мы хотим получить все элементы, похожие на образец, описываемый этим запросом.
На компьютере решить задачу ассоциативного поиска можно простым перебором с проверкой всех данных на соответствие критериям поиска. Но полный перебор трудоемок, гораздо быстрее поиск осуществляется, если данные заранее подготовлены так, что позволяют быстро формировать выборки, удовлетворяющие определенным критериям. Например, поиск в адресной книге можно значительно ускорить, если создать оглавление по первой букве.
Подготовленная должным образом память называется ассоциативной. Практическая реализация ассоциативности может использовать таблицы индексов, хэш-адресацию, бинарные деревья и тому подобные инструменты.
Независимо от реализации, ассоциативный поиск требует задания критериев похожести или близости. Как мы уже говорили, основные виды близости – это близость по описанию, близость по совместному проявлению и близость по общему явлению. Первая говорит о похожести по форме, остальные о сходстве по некой сути. Это напоминает поиск товара в Интернет-магазине. Если вы введете название, то получите список товаров, названия которых будут близки к заданному. Но кроме этого вы, скорее всего, получите еще один список, который будет называться, что-то вроде: «те, кто ищут этот товар, часто обращают внимание на…». При этом элементы второго списка могут по своим описаниям никак не походить на описание, сформулированное исходным запросом.
Посмотрим, как рассуждения об ассоциативности применимы к понятиям, используемым в нашей модели. Понятие у нас – это набор паттернов нейронов-детекторов, настроенных на один и тот же образ. Очень похоже, что одному паттерну соответствует отдельная кортикальная миниколонка. Образ – это определенное сочетание сигналов на рецептивном поле нейронов. На сенсорных зонах коры образ может быть сформирован сигналами топографической проекции. На более высоких уровнях образ – это узор активности, возникающий вокруг нейрона, при прохождении мимо него волны идентификаторов.
Для перехода нейрона в состояние вызванной активности требуется чтобы образ достаточно точно совпал с картиной весов на его синапсах. Так если нейрон настроен на детектирование определенного сочетания понятий, то волновой идентификатор должен содержать существенную часть идентификаторов каждого из них.
Срабатывание нейрона, когда повторяется большая часть понятий, составляющих его характерный стимул, называется узнаванием. Узнавание и связанная с ним вызванная активность не дают обобщенной ассоциативности. В таком режиме нейрон не реагирует ни на понятия, имеющие временную близость, ни на понятия, хотя и входящие в описание характерного для нейрона стимула, но встретившиеся отдельно от остального описания.
Нейроны, которые мы используем в нашей модели, и которые мы сравниваем с реальными нейронами мозга, способны хранить информацию не только за счет изменения синаптических весов, но и благодаря изменениям в метаботропных рецептивных кластерах. За счет последних нейроны способны запоминать фрагменты картин окружающей активности. Повторение такой запомненной картины не ведет к появлению вызванной активности нейрона, состоящей из серии импульсов, а вызывает единичный спайк. Запомнит или нет нейрон что-либо таким образом определяется рядом факторов один из которых — изменение мембранного потенциала этого нейрона. Увеличение мембранного потенциала вызывает расхождение расщепленных концов метаботропных рецепторов, что заставляет их обрести необходимую для запоминания чувствительность.
Когда нейрон узнает характерный образ и переходит в состояние вызванной активности, его мембранный потенциал существенно повышается, что дает ему возможность запоминать на внесинаптической части мембраны окружающие его картины активности.
Можно предположить, что в момент вызванной активности нейрон запоминает все окружающие его волновые узоры. Такая операция имеет вполне определенный смысл. Предположим, что мы имеем полное описание происходящего, складывающееся из большого числа понятий. Часть из них позволит нейрону узнать то понятие, детектором которого является его паттерн (миниколонка). Узнавание обеспечит этому понятию участие в общем описании. При этом на нейронах, ответственных за само понятие зафиксируется, какие еще понятия входили в полную описательную картину.
Эта фиксация произойдет на всех нейронах, составляющих наш детекторный паттерн (миниколонку). Регулярное повторение такой процедуры приведет к накоплению на поверхности нейронов-детекторов рецептивных кластеров, чувствительных к определенным сочетаниям идентификаторов.
Можно предположить, что чем чаще какое-либо понятие будет встречаться совместно с активностью детекторного паттерна, тем выше будет вероятность того, что его самостоятельное появление будет определяться внесинаптическими рецепторами и вызывать единичный спайк. То есть если между понятиями присутствует совместность в проявлении, то нейроны одного понятия будут реагировать одиночным спайком, когда в волновом описании будет появляться другое ассоциативно связанное с ним понятие.
Одновременный одиночный спайк нейронов одного детекторного паттерна – это основание для запуска соответствующей идентификационной волны. Получается, что задав одно понятие и распространив его волновой идентификатор можно получить в волновом описании набор понятий, ассоциативно с ним связанных. Это можно сопоставить с тем, как база данных по запросу выдает список всех записей, попадающих под критерии этого запроса. Получив список от базы данных, мы используем дополнительные алгоритмы, чтобы продолжить работу с этим списком. Аналогично можно представить и работу мозга. Получение доступа к ассоциациям – это только один из этапов его работы. На активность нейронов можно наложить дополнительные ограничения, которые позволят только части ассоциаций пробиться в волновое описание. Суть этих дополнительных ограничений во многом и определяет информационные процедуры, свойственные мышлению.
Пространственная самоорганизация
Мы исходим из того, что явления внешнего мира воздействуют на наши органы чувств, вызывая определенные потоки сигналов в нервных клетках. В процессе обучения кора приобретает способность детектировать определенные сочетания сигналов. Детекторами выступают нейроны, синаптические веса которых настраиваются на картины активности, соответствующие детектируемым явлениям. Нейроны коры следят за своим локальным окружением, образующим их рецептивное поле. Информация на рецептивные поля нейронов поступает либо посредством топографической проекции, либо через распространение волн идентификаторов, несущих уникальные узоры, соответствующие уже выделенным признакам. Нейроны-детекторы, реагирующие на одно и то же сочетание признаков, образуют детекторные паттерны. Узоры этих паттернов определяют уникальные волны идентификаторов, которые эти паттерны запускают, приходя в состояние вызванной активности.
В 1952 году Алан Тьюринг опубликовал работу под названием «Химические основы морфогенеза» (Turing A. M., 1952), посвященную самоорганизации материи. Сформулированный им основной принцип гласил, что глобальный порядок определяется локальным взаимодействием. То есть, чтобы получить структурную организацию всей системы, необязательно иметь некий глобальный план, а можно ограничиться исключительно заданием правил близкого взаимодействия образующих систему элементов.
Обучение нейронов происходит не изолированно, а с учетом активности их окружения. Правила учета этой активности определяют самоорганизацию коры. Самоорганизация означает, что по ходу обучения не просто формируются детектирующие нейронные паттерны, а что эти паттерны выстраиваются в некие пространственные конструкции, имеющие свой определенный смысл.
Наиболее очевидный способ самоорганизации – это структурирование по степени близости. Можно обучать нейроны так, чтобы паттерны, соответствующие близким в некотором смысле понятиям, оказывались рядом и в пространстве коры. Позже мы увидим, что такое размещение окажется необходимым для реализации многих важных функций, свойственных мозгу. Пока же просто посмотрим на механизмы, которые могут обеспечить такую организацию.
Достаточно неоднозначный вопрос – как мерить близость понятий. Один подход связан с тем, что любое понятие можно сопоставить с неким описанием. Например, описанием события может быть вектор, компоненты которого показывают выраженность в событии определенных признаков. Набор признаков, в которых ведется описание, образует описательный базис. В таком случае, разумная мера близости – это близость описаний. Чем ближе описание двух явлений, тем более похожи друг на друга эти явления. В зависимости от задачи, которая далее решается с использованием меры близости, можно выбрать тот или иной алгоритм расчета. Мера близости тесно связана с понятием расстояния между объектами. Одно можно пересчитать в другое и наоборот.
Иной подход – это временная близость явлений. Если два явления происходят одновременно, но при этом имеют разные описания, то все равно можно говорить об их определенной близости. Чем чаще явления случаются совместно, тем сильнее может быть истолкована их близость.
На самом деле эти два подхода не противоречат, а взаимодополняют друг друга. Если два явления часто случаются совместно, то надо сделать вывод о наличии явления, которое является реализацией этой совместности. Это новое явление само может быть признаком, описывающим оба исходных явления. Теперь у этих явлений появилось общее в описании, а значит, и близость в описательном подходе. Будем называть близость, учитывающую одновременно и описательную и временную близость, обобщенной близостью.
Представьте, что вы разглядываете какой-либо предмет с разных сторон. Возможно, эти виды будут совершенно различны в описательном плане, но тем не менее нельзя отрицать их близость в некотором ином смысле. Временная близость – это возможное проявление этого иного смысла. Несколько позже мы значительно глубже разовьем эту тему, сейчас же просто отметим необходимость учитывать оба подхода при определении близости понятий.
Наиболее простая для понимания нейронная сеть, которая хорошо иллюстрирует самоорганизацию по степени описательной близости – это самоорганизующиеся карты Кохонена (рисунок ниже).
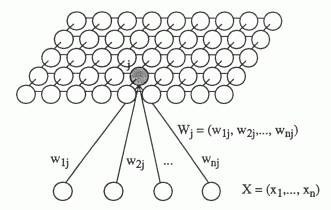
Пусть у нас есть входная информация, заданная вектором \(x\). Есть двумерная решетка из нейронов. Каждый нейрон связан с входным вектором \(x\), эта связь определяется набором весов \(\omega _{j}\). Вначале инициируем сеть случайными малыми весами. Подавая входной сигнал, для каждого нейрона можно определить его уровень активности как линейного сумматора. Возьмем нейрон, который покажет наибольшую активность, и назовем его нейроном-победителем. Далее сдвинем его веса в сторону образа, на который он оказался похож. Более того, проделаем аналогичную процедуру для всех его соседей. Будем ослаблять этот сдвиг по мере удаления от нейрона-победителя.
\[\omega _{j}(n+1)=\omega _{j}(n)+\eta(n) h_{j,i(x)}(n)(x-\omega _{j}(n))\]
Здесь – \(\eta(n)\) скорость обучения, которая падает со временем,\(h_{j,i(x)}(n)\) — амплитуда топологической окрестности (зависимость от предполагает, что она тоже со временем уменьшается).
Амплитуда окрестности может быть выбрана, например, Гауссовой функцией:
\[h_{j,i(x)}(n)=exp\left ( -\frac{d_{j,i}^{2}}{2\sigma^{2}(n) } \right )\]
Где \(d\) – расстояние между корректируемым нейроном \(j\) и нейроном-победителем \(i\).
Функция Гаусса
По мере обучения на такой самоорганизующейся карте будут выделяться зоны, соответствующие тому, как распределены обучающие образы. То есть сеть сама определит, когда во входном потоке встретятся похожие друг на друга картины, и создаст для них близкие представительства на карте. При этом, чем сильнее будут различаться образы, тем более обособленно друг от друга будут расположены и их представительства. В итоге, если соответствующим образом раскрасить результат обучения, то он будет выглядеть примерно так, как показано на рисунке ниже.

Результат обучения карты Коханена
Карты Коханена используют функцию амплитуды топологической окрестности, что предполагает, что нейроны кроме взаимодействия через синапсы могут обмениваться дополнительной информацией, говорящей о характере окружающей активности, и эта информация может влиять на ход их синаптического обучения. Необходимость в передаче дополнительной информации возникала несколько раньше и в нашей модели. Описывая обучение внесинаптических рецепторов, мы вводили правила, опирающиеся на знание нейронами окружающей активности определенного типа. Например, знание общего уровня активности позволяло нам принимать решения как о необходимости обучения, так и об отказе от него.
Чтобы оставаться при построении модели в рамках определенной биологической достоверности, попробуем показать, какие механизмы в реальной коре могут отвечать за расчет и передачу дополнительной незакодированной в сигналы аксонов информации.
Около 40 процентов объема мозга занимают глиальные клетки. Их общее количество где-то на порядок превосходит количество нейронов. Традиционно на глиальные клетки возлагается масса сервисных функций. Они создают объемный каркас, заполняя пространство между нейронами. Участвуют в поддержании гомеостаза среды. В процессе развития нервной системы задействованы для формирования топологии мозга. Шванновские клетки и олигодендроциты отвечают за миелинизацию крупных аксонов, что приводит к многократному ускорению передачи нервных импульсов.
Так как глиальные клетки не генерируют потенциалов действия, они не участвуют напрямую в информационном взаимодействии. Но это не значит, что они полностью лишены информационных функций. Например, плазматические астроциты располагаются в сером веществе и имеют многочисленные сильно ветвящиеся отростки. Эти отростки опоясывают окружающие синапсы и влияют на их работу (рисунок ниже).
Астроцит и синапс (Филдз, 2004)
Так, например, был описан следующий механизм (R. D. Fields, B. Stevens-Graham, 2002). Активация нейрона ведет к тому, что из его аксона высвобождаются молекулы АТФ. АТФ (аденозинтрифосфат) — нуклеотид, играющий исключительно важную роль во всем организме, основная его функция – обеспечение энергетических процессов. Но кроме этого АТФ способен выступать и как сигнальное вещество. Под его воздействием инициируется перемещение кальция внутрь астроцита. Это, в свою очередь, приводит к тому, что астроцит высвобождает собственный АТФ. В результате происходит передача такого состояния на соседние астроциты, которые передают его еще дальше. При этом поглощение кальция астроцитом ведет к тому, что он начинает влиять на синапсы, с которыми контактирует. Астроциты способны как усиливать реакцию синапса за счет выброса соответствующего медиатора, так и ослаблять ее за счет его поглощения или выброса связывающих нейромедиатор белков. Кроме того, астроциты способны выделять сигнальные молекулы, регулирующие выброс медиатора аксоном. Концепция передачи сигналов между нейронами, учитывающая влияние астроцитов, называется трехсторонним синапсом.
Такое взаимодействие астроцитов и нейронов не передает конкретных информационных образов, но оно очень хорошо подходит на роль механизма, обеспечивающего распространение «поля активности», которое может управлять обучением синапсов и таким образом задавать пространственные координаты для новых детекторных паттернов.
Кроме астроцитов на поведение синапсов влияет еще и межклеточный матрикс. Матрикс – это множество молекул, произведенных клетками мозга и заполняющих межклеточное пространство. В статье (Dityatev A., Schachner M., Sonderegger P., 2010) показано, что изменение состава матрикса влияет на характер синаптической пластичности, то есть на обучение нейронов.
Импульсная активность нейронов создает точечный пространственный узор, который динамично меняется, кодируя информационные потоки. Эта активность изменяет состояние окружающей глиальной среды и среды матрикса таким образом, что создается нечто наподобие поля обобщенной активности (рисунок ниже).

Точечная активность и поле активности
Поле активности, с одной стороны, размывает точечную активность, создавая область, простирающуюся за пределы активных нейронов, с другой стороны, обладает инерционностью и продолжает существовать какое-то время после прекращения импульсной активности.
Если внутри этого поля активности мы создадим детектор текущего подаваемого образа, то он окажется поблизости от похожих на него детекторов. При этом похожесть может оказаться как похожестью по рецептивному полю, так и похожестью, возникающей за счет совмещения событий по времени.
В реальном мозге лучше всего пространственная организация изучена для зоны первичной зрительной коры. За счет преобразований, начинающихся еще в сетчатке, на первичную кору поступает сигнал, в котором основная информация – это линии, описывающие контуры объектов на исходном изображении. Нейроны первичной зрительной коры видят, в основном, небольшие фрагменты этих линий, проходящие через их рецептивные поля. Неудивительно, что существенная часть нейронов этой зоны – это детекторы линий, идущих под различными углами.
Экспериментально выявлено, что нейроны, располагающиеся вертикально друг под другом, реагируют на один и тот же стимул. Группу таких нейронов принято называть кортикальной мини колонкой. Вернон Маунткасл (В. Маунткасл, Дж. Эдельман, 1981) выдвинул гипотезу, что для мозга кортикальная колонка – это основная структурная единица переработки информации.
Мы ранее, говоря о паттернах нейронов детекторов, изображали их группами нейронов, распределенных в некой локальной области. Это было следствием того, что при моделировании, и соответственно, подготовке картинок использовались плоские нейронные сети. Реальная кора трехмерна. Объемность коры не влияет на наши рассуждения о возникновении и распространении волн идентификаторов. В трехмерной коре волны распространяются точно также как и в плоской. Но в объемной коре ничто не мешает нам, расположить нейроны-детекторы, образующие единый паттерн, вертикально друг под другом. Такое расположение ни хуже и не лучше любых других. Главное требование к паттерну – это случайность его узора. Так как связи нейронов распределены случайным образом, то расположенные вертикально в одной кортикальной колонке нейроны вполне можно считать случайным паттерном. Такое вертикальное расположение достаточно удобно при определении места создания детекторного паттерна. Не надо определять локальную область, а достаточно указать позицию, в которой требуется создать паттерн. Выбор позиции может быть, например, там, где поле активности максимально для всех свободных колонок некой окрестности. Можно предположить, что кортикальные миниколонки реальной коры – это и есть детекторные паттерны, описываемые в нашей модели.
Для традиционных моделей коры, не учитывающих волновые сигналы, объяснение того, что все нейроны мини колонки реагируют на один и тот же стимул, представляет определенное затруднение. Приходится предполагать, что это либо дублирование для обеспечения отказоустойчивости, либо попытка собрать вместе нейроны, реагирующие на один стимул, но настроенные на его определение в разных позициях общего рецептивного поля. Последнее обыгрывалось в неокогнитроне через использование плоскостей простых клеток. Волновая модель позволяет взглянуть на это с иной стороны. Можно предположить, что задача кортикальной мини колонки – это, узнав характерный стимул, запустить соответствующий волновой идентификатор. Тогда несколько десятков нейронов, образующих мини колонку и реагирующих совместно, — это и есть механизм запуска волны, то есть детекторный паттерн.
Организация ориентационных колонок в реальной зрительной коре образует так называемые «волчки» («pinwheel»). Такой «волчок» (рисунок ниже (B)) имеет центр, куда сходятся колонки разных ориентаций, и расходящиеся хвосты, для которых характерна плавность смены предпочитаемого стимула. Один волчок образует гиперколонку. Хвосты разных гиперколонок переходят друг в друга, образуя ориентационную карту коры (рисунок ниже (A)).

Распределение ориентационных колонок в реальной коре, полученное оптическим методом (Николлс Дж., Мартин Р., Валлас Б., Фукс П., 2003)
На примере зрительной коры удобно сравнить реальное распределение ориентационных колонок и результаты моделирования с использованием поля активности.
Будем подавать на фрагмент коры линии под различными углами. Активность нейронов-детекторов будем считать, как косинус угла между поданным образом и образом на детекторе.
\[A_{j}=cos(\varphi)\]
Нейрон с индексом \(j\) имеет координаты на коре \((x_{j}, y_{j})\). Значение поля активности \(f_{j}\)от нейрона \(j\) в точке с координатами \(x,y\) можно представить, например, через Гауссово распределение:
\[f_{j}(x,y)=A_{j}\star exp\left ( -\frac{(x_{j}-x)^{2}+(y_{j}-y)^{2}}{2\sigma ^{2}} \right )\]
В каждой точке коры итоговое значение поля активности можно записать:
\[f(x,y)=\sum_{j}f_{j}(x,y)\]
Будем размещать новые детекторы в такой свободной позиции, для которой значение максимально. Результат такого обучения показан на рисунке ниже.

Результат обучения детекторов (слева), поле активности (справа)
Как видно, результат моделирования очень похож на реальное распределение. Но надо отметить, что приведенный пример очень условен. В реальной коре каждая колонка несколько смещена относительно соседей и следит за немного другим фрагментом изображения. Это создает «хвосты» из детекторов, настроенных на одну ориентацию. Но при этом стимулы таких детекторов – это разные образы, основанные на разных рецептивных полях.
Пространственную самоорганизацию коры можно сопоставить с самоорганизацией материи в окружающем нас мире. Атомы, молекулы, предметы, планеты, звезды, галактики, вселенная – все это следствие существования четырех фундаментальных взаимодействий. Принято говорить о гравитационном, электромагнитном, сильном и слабом взаимодействии. Частицы материи создают вокруг себя поля, соответствующие взаимодействиям. Однотипные поля суммируются и формируют результирующее поле. Результирующие поля оказывают влияние на частицы, определяя их поведение. Где-то так же обстоит дело и с мозгом. Похоже, что кора может создавать несколько типов полей, обладающих разными свойствами. Каждое из полей оказывает свое воздействие на поведение нейронов. Совокупность этих взаимодействий, например, может определять алгоритм обучения синапсов и тем самым пространственную организацию нейронов-детекторов.
Нетрудно заметить, что пространственная группировка по обобщенной близости содержит внутреннее противоречие. В связи с этим вспоминается анекдот. Решил как-то лев поделить зверей на красивых и умных. И вот стоит растерянная обезьяна: «И что мне теперь, разорваться?». Сложности возникают, когда размещаемое понятие оказывается близко к другим, удаленным друг от друга понятиям. Например, если мы раскладываем книги по полкам библиотеки, то нам одновременно хочется сгруппировать их и по жанрам, и по авторам, и по языку оригинала, и по читательскому рейтингу. Но если книга только одна, то возникает определенная проблема.
В процессе самоорганизации коры может оказаться, что одно и то же понятие близко в обобщенном смысле сразу к нескольким другим, расположенным в разных местах коры. Не имеет смысла размещать детекторный паттерн компромиссно, пытаясь найти некое равноудаленное место. Во-первых, такого места может не быть, а во-вторых, это полностью нарушит принцип «похожее рядом». Не остается ничего другого, как создать несколько детекторных паттернов, каждый поблизости от своего локального максимума поля активности.
На первый взгляд, дублирование одного и того же понятия в разных местах выглядит некрасиво. Работая с информацией, всегда хочется прийти к ее интеграции, а не распылению. Но наша модель нейронной сети позволяет достаточно изящно снять возникшие противоречия. Каждый из детекторных паттернов, хотя и создается в разных местах коры, обучается на одну и ту же волну идентификаторов. При этом, если брать двухуровневую конструкцию, один из уровней фиксирует на себе фрагмент узора волны общего идентификатора. Это означает, что разбросанные по разным местам детекторные паттерны оказываются связаны между собой в единое понятие с общим идентификатором.
Такое поведение понятий – следствие задекларированного ранее дуализма. Понятие – это одновременно и паттерн и волна. Паттерны порождают волну, волна активирует паттерны. Пространственное положение паттернов, относящихся к одному понятию, характеризует понятие через его близость к другим понятиям. Положение паттернов на коре, по сути, очень многое говорит о свойствах и характеристиках соответствующего явления. Но если в картах Кохонена одному понятию соответствует одна область, как получается из парадигмы «победитель забирает все», то в нашей модели удается не только дать более богатое описание, но за счет дуализма еще и сохранить единство составляющих.
Самоорганизация по принципу «похожее рядом» не является для коры самоцелью. Позже мы покажем, что важнейшие для работы мозга алгоритмы требуют именно такой организации и практически нереализуемы вне ее.
А теперь не совсем банальная физическая аналогия. Основная идея квантовой физики заключается в том, что квантовая система может находиться не в любых возможных состояниях, а только в некоторых, допустимых для нее. При этом квантовая система не принимает никакого определенного значения, пока не произойдет измерение. Только в результате измерения система переходит в одно из разрешенных ей состояний. Под измерением понимается любое внешнее взаимодействие, которое заставляет квантовую систему проявить себя.
В квантовой физике вводится комплекснозначная функция , описывающая чистое состояние объекта, которая называется волновой функцией. В наиболее распространенной копенгагенской интерпретации эта функция связана с вероятностью обнаружения объекта в одном из чистых состояний (квадрат модуля волновой функции представляет собой плотность вероятности). Гамильтоновой системой называется динамическая система, описывающей физические процессы без диссипации. Пока квантовая система не вступит во взаимодействие, она является гамильтоновой. Описать поведение такой системы во времени и пространстве можно, описав эволюцию ее волновой функции. Эта эволюция определяется уравнением Шредингера.
\[i\hbar\frac{\partial }{\partial t}\Psi =\widehat{H} \Psi\]
Где – \(\widehat{H}\) гамильтониан – оператор полной энергии системы. Его спектр – это множество возможных состояний, в которых система может оказаться после измерения.
Когда происходит измерение, квантовая система принимает одно из разрешенных состояний. Это называется редукцией или коллапсом волновой функции. В процессе измерения система перестает быть изолированной, и ее энергия может не сохраняться, так как происходит обмен энергией с прибором.
Какое состояние примет квантовая система в момент измерения – дело случая. Но вероятность выбора каждого из состояний не одинакова, она определяется связанным с этим состоянием значением волновой функции.
В нашей модели детекторные паттерны создаются в позициях коры, где наблюдается описательная или временная близость нового понятия и понятий уже существующих. Тем самым формируется набор мест, описывающих вероятность проявления понятия в той или иной ситуации. Это можно сравнить со спектром гамильтониана полной энергии квантовой системы. Распространение волн идентификаторов можно сопоставить с эволюцией волновой функции. Описание, которое несет идентификатор, переходит в активность паттернов. Далее мы покажем, что при этом активируются не все паттерны, а только те, что могут описать непротиворечивую картину, соответствующую контексту происходящего. В этот момент наша система переходит в одно из возможных разрешенных состояний, что сильно напоминает коллапс волновой функции.